Impact of Trematode Parasitism on the Fauna of a North Sea Tidal Flat
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Effect of Temperature on the Infectivity of Metacercariae of Zygocotyle Lunata (Digenea: Paramphistomidae)
J. Parasitol., 87(1), 2001, p. 10±13 q American Society of Parasitologists 2001 EFFECT OF TEMPERATURE ON THE INFECTIVITY OF METACERCARIAE OF ZYGOCOTYLE LUNATA (DIGENEA: PARAMPHISTOMIDAE) David L. Ferrell*, Nicholas J. Negovetich, and Eric J. Wetzel² Department of Biology, Wabash College, P.O. Box 352, Crawfordsville, Indiana 47933 ABSTRACT: As a test of the energy limitation hypothesis (ELH), we predicted that temperature would have a signi®cant in¯uence on the infectivity of metacercariae of the digenetic trematode Zygocotyle lunata. Snails infected with Z. lunata were collected from ponds near Crawfordsville, Indiana, isolated at room temperature, and examined for the release of cercariae. Newly encysted metacercariae were collected and incubated 1±30 days at 1 of 5 temperatures (0, 3, 25, 31, 37 C). Twenty-®ve cysts were fed to each of 5 or 10 mice per treatment group (temperature). At 17 days postinfection, mice were killed and worms were recovered; data were collected on levels of infection in each group and the total body area of each worm. No worms were found in mice fed cysts that had been held at0Cor37C(after 30 days). There were no differences in prevalence, infectivity, or mean intensity among the 3, 25, and 31 C treatments. Infectivity of metacercariae incubated at 37 C for 1 day was signi®cantly greater than in all other treatments, while infectivity of metacercariae in the 37 C/15-day treatment was signi®cantly lower than in all others. Mean body area of worms at 37 C/15 days was signi®cantly greater than at other temperatures, suggesting density-dependent increases in growth. -

Parasite Infection of the Non-Indigenous Round Goby (Neogobius Melanostomus) in the Baltic Sea
Downloaded from orbit.dtu.dk on: Oct 04, 2021 Parasite infection of the non-indigenous round goby (Neogobius melanostomus) in the Baltic Sea Ojaveer, Henn; Turovski, Aleksei; Nõomaa, Kristiina Published in: Aquatic Invasions Publication date: 2020 Document Version Peer reviewed version Link back to DTU Orbit Citation (APA): Ojaveer, H., Turovski, A., & Nõomaa, K. (2020). Parasite infection of the non-indigenous round goby (Neogobius melanostomus) in the Baltic Sea. Aquatic Invasions, 15(1), 160-176. General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Users may download and print one copy of any publication from the public portal for the purpose of private study or research. You may not further distribute the material or use it for any profit-making activity or commercial gain You may freely distribute the URL identifying the publication in the public portal If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Aquatic Invasions (2020) Volume 15 Article in press Special Issue: Proceedings of the 10th International Conference on Marine Bioinvasions Guest editors: Amy Fowler, April Blakeslee, Carolyn Tepolt, Alejandro Bortolus, Evangelina Schwindt and Joana Dias CORRECTED PROOF Research Article Parasite infection of the non-indigenous round goby (Neogobius melanostomus) in the Baltic Sea Henn Ojaveer1,2,*, Aleksei Turovski3 and Kristiina Nõomaa4 1University of Tartu, Ringi 35, 80012 Pärnu, Estonia 2National Institute of Aquatic Resources, Technical University of Denmark, Kemitorvet Building 201, 2800 Kgs. -

Periwinkles Littorina Littorea Sampled Close to Charr Farms in Northern Norway
DISEASES OF AQUATIC ORGANISMS Vol. 12: 59-65, 1991 Published December 5 Dis. aquat. Org. l Occurrence of the digenean Cryptocotyle lingua in farmed Arctic charr Salvelinus alpinus and periwinkles Littorina littorea sampled close to charr farms in northern Norway Roar Kristoffersen Department of Aquatic Biology, Norwegian College of Fishery Science, University of Trornso, Dramsveien 201B, N-9000 Tromso, Norway ABSTRACT: Occurrence of Cryptocotyle lingua rediae was recorded in periwinkle samples collected adjacent to 10 charr farms and at control sites 1 to 5 km from the farms. In 7 out of 10 localities the prevalence of infection was higher in the sample taken adjacent to the farm than in the control, and overall prevalence was 13.7 O/O in periwinkles near the farms and 6.1 % in snails from the control sites, a highly significant difference. Prevalences in periwinkles close to farms tended to increase with duration of farming at the site. The role of the final host, piscivorous birds, is considered Samples of charr from 11 farms were investigated for visible black spots caused by encysted C. llngua metacercariae. No infected charr were recorded in the 2 land-based farms where seawater exposed to UV-light (photozone) was pumped to the tanks, whilst 83.2 '10 of the fish exhibited black spots in the 9 farms where the charr were stocked in floating net cages in the sea. In most infected fish the C. ljngua cysts were located only on the fins in relatively small numbers. INTRODUCTION transmission from such focal points to wild host popula- tions and vice versa. -

First Characterisation of the Populations and Immune-Related
First characterisation of the populations and immune-related activities of hemocytes from two edible gastropod species, the disk abalone, Haliotis discus discus and the spiny top shell, Turbo cornutus. Ludovic Donaghy, Hyun-Ki Hong, Christophe Lambert, Heung-Sik Park, Won Joon Shim, Kwang-Sik Choi To cite this version: Ludovic Donaghy, Hyun-Ki Hong, Christophe Lambert, Heung-Sik Park, Won Joon Shim, et al.. First characterisation of the populations and immune-related activities of hemocytes from two edible gastropod species, the disk abalone, Haliotis discus discus and the spiny top shell, Turbo cornutus.. Fish and Shellfish Immunology, Elsevier, 2010, 28 (1), pp.87-97. 10.1016/j.fsi.2009.10.006. hal- 00460531 HAL Id: hal-00460531 https://hal.archives-ouvertes.fr/hal-00460531 Submitted on 1 Mar 2010 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. First characterisation of the populations and immune-related activities of hemocytes from two edible gastropod species, the disk abalone, Haliotis discus discus and the spiny top shell, Turbo cornutus . Ludovic Donaghy a,b,* , Hyun-Ki Hong a, Christophe Lambert b, Heung-Sik Park c, Won Joon Shim d, Kwang-Sik Choi a. -

Behavioral Analysis of a Digenetic Trematode Cercaria (Microphallus Turgidus) in Relation to the Microhabitat of Grass Shrimp (Palaemonetes Spp.)
Georgia Southern University Digital Commons@Georgia Southern Electronic Theses and Dissertations Graduate Studies, Jack N. Averitt College of Fall 2010 Behavioral Analysis of a Digenetic Trematode Cercaria (Microphallus Turgidus) in Relation to The Microhabitat of Grass Shrimp (Palaemonetes Spp.) Patricia O'Leary Follow this and additional works at: https://digitalcommons.georgiasouthern.edu/etd Recommended Citation O'Leary, Patricia, "Behavioral Analysis of a Digenetic Trematode Cercaria (Microphallus Turgidus) in Relation to The Microhabitat of Grass Shrimp (Palaemonetes Spp.)" (2010). Electronic Theses and Dissertations. 743. https://digitalcommons.georgiasouthern.edu/etd/743 This thesis (open access) is brought to you for free and open access by the Graduate Studies, Jack N. Averitt College of at Digital Commons@Georgia Southern. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of Digital Commons@Georgia Southern. For more information, please contact [email protected]. Behavioral analysis of a digenetic trematode cercaria ( Microphallus turgidus ) in relation to the microhabitat of grass shrimp ( Palaemonetes spp.) by Patricia O’Leary (Under the Direction of Oscar J. Pung) Abstract The hydrobiid snail and grass shrimp hosts of the microphallid trematode Microphallus turgidus are found in specific microhabitats. The primary second intermediate host of this parasite is the grass shrimp Palaemonetes pugio. The behavior of trematode cercaria often reflects the habitat and behavior of the host species. The objective of my study was to examine the behavior of M. turgidus in relation to the microhabitat selection of the second intermediate host. To do so, I established a behavioral ethogram for the cercariae of M. turgidus and compared the behavior of these parasites to the known host behavior. -

MOLLUSCS Species Names – for Consultation 1
MOLLUSCS species names – for consultation English name ‘Standard’ Gaelic name Gen Scientific name Notes Neologisms in italics der MOLLUSC moileasg m MOLLUSCS moileasgan SEASHELL slige mhara f SEASHELLS sligean mara SHELLFISH (singular) maorach m SHELLFISH (plural) maoraich UNIVALVE SHELLFISH aon-mhogalach m (singular) UNIVALVE SHELLFISH aon-mhogalaich (plural) BIVALVE SHELLFISH dà-mhogalach m (singular) BIVALVE SHELLFISH dà-mhogalaich (plural) LIMPET (general) bàirneach f LIMPETS bàirnich common limpet bàirneach chumanta f Patella vulgata ‘common limpet’ slit limpet bàirneach eagach f Emarginula fissura ‘notched limpet’ keyhole limpet bàirneach thollta f Diodora graeca ‘holed limpet’ china limpet bàirneach dhromanach f Patella ulyssiponensis ‘ridged limpet’ blue-rayed limpet copan Moire m Patella pellucida ‘The Virgin Mary’s cup’ tortoiseshell limpet bàirneach riabhach f Testudinalia ‘brindled limpet’ testudinalis white tortoiseshell bàirneach bhàn f Tectura virginea ‘fair limpet’ limpet TOP SHELL brùiteag f TOP SHELLS brùiteagan f painted top brùiteag dhotamain f Calliostoma ‘spinning top shell’ zizyphinum turban top brùiteag thurbain f Gibbula magus ‘turban top shell’ grey top brùiteag liath f Gibbula cineraria ‘grey top shell’ flat top brùiteag thollta f Gibbula umbilicalis ‘holed top shell’ pheasant shell slige easaig f Tricolia pullus ‘pheasant shell’ WINKLE (general) faochag f WINKLES faochagan f banded chink shell faochag chlaiseach bhannach f Lacuna vincta ‘banded grooved winkle’ common winkle faochag chumanta f Littorina littorea ‘common winkle’ rough winkle (group) faochag gharbh f Littorina spp. ‘rough winkle’ small winkle faochag bheag f Melarhaphe neritoides ‘small winkle’ flat winkle (2 species) faochag rèidh f Littorina mariae & L. ‘flat winkle’ 1 MOLLUSCS species names – for consultation littoralis mudsnail (group) seilcheag làthaich f Fam. -

Shelled Molluscs
Encyclopedia of Life Support Systems (EOLSS) Archimer http://www.ifremer.fr/docelec/ ©UNESCO-EOLSS Archive Institutionnelle de l’Ifremer Shelled Molluscs Berthou P.1, Poutiers J.M.2, Goulletquer P.1, Dao J.C.1 1 : Institut Français de Recherche pour l'Exploitation de la Mer, Plouzané, France 2 : Muséum National d’Histoire Naturelle, Paris, France Abstract: Shelled molluscs are comprised of bivalves and gastropods. They are settled mainly on the continental shelf as benthic and sedentary animals due to their heavy protective shell. They can stand a wide range of environmental conditions. They are found in the whole trophic chain and are particle feeders, herbivorous, carnivorous, and predators. Exploited mollusc species are numerous. The main groups of gastropods are the whelks, conchs, abalones, tops, and turbans; and those of bivalve species are oysters, mussels, scallops, and clams. They are mainly used for food, but also for ornamental purposes, in shellcraft industries and jewelery. Consumed species are produced by fisheries and aquaculture, the latter representing 75% of the total 11.4 millions metric tons landed worldwide in 1996. Aquaculture, which mainly concerns bivalves (oysters, scallops, and mussels) relies on the simple techniques of producing juveniles, natural spat collection, and hatchery, and the fact that many species are planktivores. Keywords: bivalves, gastropods, fisheries, aquaculture, biology, fishing gears, management To cite this chapter Berthou P., Poutiers J.M., Goulletquer P., Dao J.C., SHELLED MOLLUSCS, in FISHERIES AND AQUACULTURE, from Encyclopedia of Life Support Systems (EOLSS), Developed under the Auspices of the UNESCO, Eolss Publishers, Oxford ,UK, [http://www.eolss.net] 1 1. -

Physiological Responses to Ocean Acidification and Warming
Marine Environmental Research 130 (2017) 38e47 Contents lists available at ScienceDirect Marine Environmental Research journal homepage: www.elsevier.com/locate/marenvrev Physiological responses to ocean acidification and warming synergistically reduce condition of the common cockle Cerastoderma edule * E.Z. Ong a, b, , M. Briffa b, T. Moens a, C. Van Colen a a Ghent University, Biology Department, Marine Biology Research Group, Krijgslaan 281eS8, B 9000 Ghent, Belgium b Marine Biology & Ecology Research Centre, Plymouth University, Plymouth PL4 8AA, UK article info abstract Article history: The combined effect of ocean acidification and warming on the common cockle Cerastoderma edule was Received 4 February 2017 investigated in a fully crossed laboratory experiment. Survival of the examined adult organisms remained Received in revised form high and was not affected by elevated temperature (þ3 C) or lowered pH (À0.3 units). However, the 6 June 2017 morphometric condition index of the cockles incubated under high pCO conditions (i.e. combined Accepted 3 July 2017 2 warming and acidification) was significantly reduced after six weeks of incubation. Respiration rates Available online 5 July 2017 increased significantly under low pH, with highest rates measured under combined warm and low pH conditions. Calcification decreased significantly under low pH while clearance rates increased signifi- Keywords: Future ocean cantly under warm conditions and were generally lower in low pH treatments. The observed physio- fi Ocean acidification logical responses suggest that the reduced food intake under hypercapnia is insuf cient to support the Ocean warming higher energy requirements to compensate for the higher costs for basal maintenance and growth in Cerastoderma edule future high pCO2 waters. -
Show Ethnoactivity
U Emetic Plant Abrus precatorius (Paternoster; Rakat; Reglisse; Pois Rouge; Weesboontje; Ma Liao Tou; Jequerit; Hung Tou; Graines Reglisse; Peonia De St Tomas; Paratella; Peonia; Hint Meyankoku; Liane Reglisse; Gunchi; Rosary Pea; To-Azuki; Cain Ghe) Acacia farnesiana (Tusca; Kembang bandira; Cuji; Esponjeira; Kambang japun; Kembang nagasiri) Acalypha indica (Lelatang) Achyranthes aspera (Apamarga; Rarai; Feuilles La Fievre; Rabo De Gato; Chaff Tree; Jarongan; Santypite) Acorus calamus (Jerangau; Calomo Aromatico; Sweet Flag; Vacha; Ganoeak; Doringo; Seki-Sho; Jeringau; Bach; Djerango; Shui Ch'Ang Pu; Acoro Aromatico; Vaj; Kalmus; Calamus; Ch'Ang P'U Chiu; Agri Turki; Kalmoes; Calmus; Acore Vrai; Acorus; Sarango; Jariangau; Calamo Aromatico; Azakegeri; Kalmos; Kaliraga; Sho-Bu) Adenocalymna alliaceum Aesculus argutus Ageratum conyzoides (Rompesaraguelo; Rumput tahi ayam; Wedusan; Berokan; Aru batu; Bandotan) Ailanthus altissima (Gok Aghaji; Ch'Ou Ch'Uang Shu Ken; Ch'U; Lisan At Tair; Niwa-Urusi) Ailanthus glandsulosa (Kokar Agac) Alangium salviifolium Alchornea cordifolia Aletris farinosa (Sterwortel; Unicorne; Sternwurzel; Star Grass; Blazing Star; True Unicorn) Aleurites fordii (Jabilla Extranjera; Tung) Allamanda blanchetii Allamanda cathartica (Yellow Allamanda) Alnus rubra (Rode Els; Aune Rouge; Red Alder; Rote Erle) Alnus rugosa Alpinia malaccensis Amaranthus dubius (Zepina; Epinard Marron; Bledo) Ammi visnaga (Khellakraut; Khillah; Busnaga; Pick Toothh; Viznaga; Khaizaran; Biznaga; Anmi; Bisnaga Das Searas) Anagyris foetida (Domuzdikeni; -

Helminthes of Goby Fish of the Hryhoryivsky Estuary (Black Sea, Ukraine)
Vestnik zoologii, 36(3): 71—76, 2002 © Yu. Kvach, 2002 UDC 597.585.1 : 616.99(262.55) HELMINTHES OF GOBY FISH OF THE HRYHORYIVSKY ESTUARY (BLACK SEA, UKRAINE) Yu. Kvach Department of Zoology, Odessa University, Shampansky prov., 2, Odessa, 65058 Ukraine E-mail: [email protected] Accepted 4 September 2001 Helminthes of Goby Fish of the Hryhoryivsky Estuary (Black Sea, Ukraine). Kvach Yu. – In the paper the data about the helminthofauna of Neogobius melanostomus, N. ratan, N. fluviatilis, Mesogobius batrachocephalus, Zosterisessor ophiocephalus, and Proterorhynus marmoratus in the Hryhoryivsky Estu- ary are presented. The fauna of gobies’ helmint hes consist of 10 species: 5 trematods (Cryptocotyle concavum met., C. lingua met., Pygidiopsis genata met., Acanthostomum imbutiforme met.), Asymphylo- dora pontica, one cestoda (Proteocephalus gobiorum), 2 nematods (Streptocara crassicauda l., Dichelyne minutus), and 2 acanthocephalans (Acanthocephaloides propinquus, Telosentis exiguus). Only one of trematods species was presented by adult stage. The modern fauna of helminthes and published data are compared. The relative stability of the goby fish helminthofauna of the Estuary is mentioned. Key words: goby, helminth, infection, Hryhoryivsky Estuary. Ãåëüìèíòû áû÷êîâûõ ðûá Ãðèãîðüåâñêîãî ëèìàíà (×åðíîå ìîðå, Óêðàèíà). Êâà÷ Þ. – Èññëåäî- âàíà ãåëüìèíòîôàóíà Neogobius melanostomus, N. ratan, N. fluviatilis, Mesogobius batrachocephalus, Zosterisessor ophiocephalus è Proterorhynus marmoratus èç Ãðèãîðüåâñêîãî ëèìàíà. Ôàóíà ãåëüìèí- òîâ áû÷êîâ âêëþ÷àåò 10 âèäîâ. Èç íèõ 5 âèäîâ òðåìàòîä (Cryptocotyle lingua met., C. concavum met., Pygidiopsis genata met., Acanthostomum imbutiforme met., Asymphylodora pontica), îäèí âèä öåñ- òîä (Proteocephalus gobiorum), 2 âèäà íåìàòîä (Streptocara crassicauda l., Dichelyne minutus), 2 âèäà ñêðåáíåé (Acanthocephaloides propinquus, Telosentis exiguus). Èç ïÿòè âèäîâ òðåìàòîä òîëüêî îäèí ïðåäñòàâëåí âçðîñëîé ñòàäèåé. -

Mercury Concentration in Bivalve Molluscs
Bull Vet Inst Pulawy 59, 357-360, 2015 DOI:10.1515/bvip-2015-0053 Mercury concentration in bivalve molluscs Józef Szkoda1, Maciej Durkalec1, Agnieszka Nawrocka1, Mirosław Michalski2 1Department of Pharmacology and Toxicology, 2Department of Hygiene of Animal Origin, National Veterinary Research Institute, 24-100 Pulawy, Poland [email protected] Received: April 23, 2015 Accepted: September 4, 2015 Abstract A total of 85 mussel samples of eight species were examined. Analysis of mercury in the freeze-dried samples was carried out by atomic absorption spectrometry method using direct mercury analyser AMA 254. The analytical procedure for determination of mercury was covered by the quality assurance programme of research and participation in national and international proficiency tests. Concentrations of total mercury in all investigated samples were found to be generally low, in the range of 0.033-0.577 mg/kg of dry weight and of 0.003-0.045 mg/kg of wet weight. The results indicate that obtained levels of mercury in bivalve molluscs are not likely to pose a risk to the health of consumers. Keywords: mercury, bivalve molluscs, bioaccumulation, food safety. Introduction which may pose a threat to the health of consumers (7, 13). In Poland, the interest in marine food, or seafood, The rules for the marketing of live bivalve has increased in the recent years. Crustaceans, mostly molluscs for direct consumption and also for shrimps, squids, and lobsters, are frequently chosen processing are strictly defined in EU law regulating by consumers, but less popular oysters, cockles, all aspects of production, including environmental mussels, and clams which belong to bivalve molluscs conditions, farming, cleaning, transport, sale, are also purchased. -
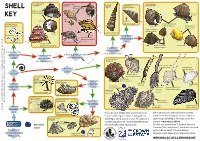
Shell Whelk Dog Whelk Turret It Could Be a Periwinkle Shell (Nucella Lapillus) Shell Spire Shell Thick Top Shell (Osilinus Lineatus) Dark Stripes Key on Body
It could be a type of It could be a type of It could be a It could be a type of topshell whelk Dog whelk turret It could be a periwinkle Shell (Nucella lapillus) shell spire shell Thick top shell (Osilinus lineatus) Dark stripes Key on body Egg Underside capsules Actual size It could be a type of (Hydrobia sp) Common periwinkle spiral worm White ‘Colar’ (Littorina littorea) Flat periwinkle (Littorinasp) Yes Roughly ‘ribbed’ shell. Very high up shore ‘Tooth inside (Turitella communis) opening (Spirorbis sp) Does it have 6 Common whelk No (Buccinum undatum) Yes or more whorls Brown, speckled Netted dog whelk body (twists)? Painted topshell (Nassarius reticulatus) (Calliostoma zizyphinum) No Rough periwinkle Flattened spire Yes Is it long, thin (Littorina saxatilis) Yes Yes and cone shaped Is it permanently No like a unicorn’s horn? attached to Is there a groove or teeth No Is there mother No a surface? in the shell opening? of pearl inside It could be a type of the shell opening? bivalve Yes Yes Common otter-shell (Lutraria lutraria) Bean-like tellin No Is the shell in (Fabulina fabula) Is it 2 parts? spiraled? Common cockle (Cerastoderma edule) It could be a Flat, rounded No sand No Great scallop mason It could be a Is the shell a (Pecten maximus) shell Razor shell worm keel worm Wedge-shaped Is the case dome or (Ensis sp) No Pacific oyster shell made from Yes cone shape? (Crassostrea gigas) Shell can be Peppery furrow shell very large (Scrobicularia plana) sand grains? Elongated and and doesn’t (Lanice conchilega) deep-bodied fully close with large ‘frills’ No (Pomatoceros sp) Yes It could be a type of sea urchin It could be a type of An acorn Native oyster Empty barnacle barnacle Does it have that may be found in estuaries and shores in the UK.