Strategies to Improve Nerve Regeneration After Radical Prostatectomy: a Narrative Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Some Aspects of Facial Nerve Paralysis* PART Li
13 Januarie 1973 S.-A. MEDIESE TVDSKRIF 65 Some Aspects of Facial Nerve Paralysis* PART lI. ELECTRICAL TESTS AND THE SUBMAXILLARY SALlYARY TEST M. G. POTGIETER,t M.B. CH.B. UNIV. PRET., M.MED. OTOL. UNIV. CAPE TOWN, Department of Otolaryngology, Groote Schuur Hospital, Observatory, Cape SUMMARY fibres are the first to be affected and when there is re generation, the new fibres conduct slowly.' The submaxillary salivary flow test gives reliable in The maintenance of nerve and muscle excitability de formation as to whether neurapraxia, axono:mesis, or pends greatly upon sodium and potassium cations. States neurotmesis of the facial nerve is present. This can be of polarization and depolarization are dependent on the corroborated by electrical studies. This test can make an shift of sodium and potassium cations across the cell important contribution to the topognosis and prognosis of membranes. With injury to the nerve;" elimination of facial paralysis, especially when elaborate electrical the membrane potential takes place and this causes a lack equipment for nerve excitability tests and electromyo of electrical conduction. A shift of potassium from the graphy is not available. cell takes place. An over-all increase in the potassium content of the injured nerve exists where, for instance, S. Afr. Med. J., 47, 65 (1973). bone fragments cause a foreign body reaction. Hyaluronic acid from an inflammatory response, occurs as a potas sium hyaluronate. Fibroblast and mast-cell activities in ELECTRlCAL TESTS crease and elaborate mucopolysaccharides. This may explain the high potassium values, maintained in the In 1872 Duchenne described the technique of nerve ex injured area of the nerve, inhibiting transmission. -
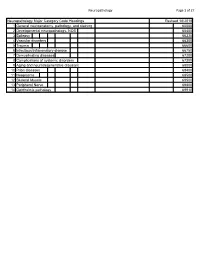
Neuropathology Category Code List
Neuropathology Page 1 of 27 Neuropathology Major Category Code Headings Revised 10/2018 1 General neuroanatomy, pathology, and staining 65000 2 Developmental neuropathology, NOS 65400 3 Epilepsy 66230 4 Vascular disorders 66300 5 Trauma 66600 6 Infectious/inflammatory disease 66750 7 Demyelinating diseases 67200 8 Complications of systemic disorders 67300 9 Aging and neurodegenerative diseases 68000 10 Prion diseases 68400 11 Neoplasms 68500 12 Skeletal Muscle 69500 13 Peripheral Nerve 69800 14 Ophthalmic pathology 69910 Neuropathology Page 2 of 27 Neuropathology 1 General neuroanatomy, pathology, and staining 65000 A Neuroanatomy, NOS 65010 1 Neocortex 65011 2 White matter 65012 3 Entorhinal cortex/hippocampus 65013 4 Deep (basal) nuclei 65014 5 Brain stem 65015 6 Cerebellum 65016 7 Spinal cord 65017 8 Pituitary 65018 9 Pineal 65019 10 Tracts 65020 11 Vascular supply 65021 12 Notochord 65022 B Cell types 65030 1 Neurons 65031 2 Astrocytes 65032 3 Oligodendroglia 65033 4 Ependyma 65034 5 Microglia and mononuclear cells 65035 6 Choroid plexus 65036 7 Meninges 65037 8 Blood vessels 65038 C Cerebrospinal fluid 65045 D Pathologic responses in neurons and axons 65050 1 Axonal degeneration/spheroid/reaction 65051 2 Central chromatolysis 65052 3 Tract degeneration 65053 4 Swollen/ballooned neurons 65054 5 Trans-synaptic neuronal degeneration 65055 6 Olivary hypertrophy 65056 7 Acute ischemic (hypoxic) cell change 65057 8 Apoptosis 65058 9 Protein aggregation 65059 10 Protein degradation/ubiquitin pathway 65060 E Neuronal nuclear inclusions 65100 -

OIICS Manual 2012
SECTION 2 Definitions, Rules of Selection, and Titles and Descriptions SECTION CONTENTS 2.1 Nature of Injury or Illness 2.2 Part of Body Affected 2.3 Source and Secondary Source of Injury or Illness 2.4 Event or Exposure *-Asterisks denote a summary level code not assigned to individual cases. _____________________________________________________________________________________________ 01/12 6 SECTION 2.1 Nature of Injury or Illness SECTION CONTENTS 2.1.1 Definition, Rules of Selection 2.1.2 Titles and Descriptions *-Asterisks denote a summary level code not assigned to individual cases. _____________________________________________________________________________________________ 01/12 7 2.1.1 Nature of Injury or Illness—Definition, Rules of Selection 1.0 DEFINITION The nature of injury or illness identifies the principal physical characteristic(s) of the work related injury or illness. RULES OF SELECTION: 1.1 Name the injury or illness indicated on the source document. Example: For strained back, choose Strains. 1.2 When two or more injuries or illnesses are indicated, and one is a sequela, aftereffect, complication due to medical treatment, or re-injury, choose the initial injury or illness. Example: If a laceration became infected developing into septicemia, choose Cuts, lacerations. 1.3 When two or more injuries or illnesses are indicated and one is more severe than the other(s) and is not a sequela or complication of the other injury or illness, select the more severe injury or illness. Example: For sprained finger and fractured wrist, choose Fractures. 1.3.1 When a single event or exposure produces an injury and transmits a disease simultaneously, and one is more severe than the other(s), select the more severe injury or disease. -

Nerve Evaluation Protocol 2014
Nerve Evaluation Protocol 2014 TABLE OF CONTENTS INTRODUCTION .................................................................................................... 1 A REVIEW OF SENSORY NERVE INJURY ................................................................ 3 TERMINOLOGY ...................................................................................................... 5 INFORMED CONSENT ............................................................................................ 6 PREOPERATIVE EVALUATION ................................................................................ 7 TESTS FOR SENSORY NERVE FUNCTION: ........................................................... 12 MATERIALS NEEDED FOR TESTING SENSORY PERCEPTION .............................. 17 TESTING TECHNIQUE .......................................................................................... 20 REFERENCES .......................................................................................................... 23 BIBLIOGRAPHY - CORONECTOMY ..................................................................... 28 SAMPLE SENSORY RECORDING SHEETS ............................................................. 30 A HANDOUT FOR PATIENTS ............................................................................... 33 INTRODUCTION The first edition of this document was produced in the Spring of 1988. Dr. A. F. Steunenberg and Dr. M. Anthony Pogrel collaborated to produce the first edition with input from Mr. Art Curley, Esquire, and with Dr. Charles Alling editing -

Novel Cryoneurolysis Device for the Treatment of Sensory and Motor Peripheral Nerves
UC San Diego UC San Diego Previously Published Works Title Novel cryoneurolysis device for the treatment of sensory and motor peripheral nerves. Permalink https://escholarship.org/uc/item/7s25k7xt Journal Expert review of medical devices, 13(8) ISSN 1743-4440 Authors Ilfeld, Brian M Preciado, Jessica Trescot, Andrea M Publication Date 2016-08-01 DOI 10.1080/17434440.2016.1204229 Peer reviewed eScholarship.org Powered by the California Digital Library University of California EXPERT REVIEW OF MEDICAL DEVICES, 2016 VOL. 13, NO. 8, 713–725 http://dx.doi.org/10.1080/17434440.2016.1204229 REVIEW Novel cryoneurolysis device for the treatment of sensory and motor peripheral nerves Brian M. Ilfelda, Jessica Preciadob and Andrea M. Trescotc aDepartment of Anesthesiology, University California San Diego, San Diego, CA, USA; bMyoscience, Inc., Fremont, CA, USA; cAlaska Pain Management Center, Wasilla, AK, USA ABSTRACT ARTICLE HISTORY Introduction: Cryoneurolysis is the direct application of low temperatures to reversibly ablate periph- Received 5 January 2016 eral nerves to provide pain relief. Recent development of a handheld cryoneurolysis device with small Accepted 17 June 2016 gauge probes and an integrated skin warmer broadens the clinical applications to include treatment of Published online superficial nerves, further enabling treatments for pre-operative pain, post-surgical pain, chronic pain, 13 July 2016 and muscle movement disorders. KEYWORDS Areas covered: Cryoneurolysis is the direct application of cold temperatures to a peripheral nerve, Cryoneurolysis; resulting in reversible ablation due to Wallerian degeneration and nerve regeneration. Use over the last cryoanalgesia; 50 years attests to a very low incidence of complications and adverse effects. -

Autophagy in Peripheral Neuropathy
Linköping University Medical Dissertation No. 1582 Autophagy in Peripheral Neuropathy Ayman Osman Department of Clinical and Experimental Medicine Linköping University, Sweden Linköping 2017 Autophagy in Peripheral Neuropathy Ayman Osman, 2017 Cover/picture/Illustration/Design: The cover picture by Kristin Samuelsson from Karolinska institute. All electron microscope pictures by Simin Mohseni and Ayman Osman. Confocal microscope pictures by Ayman Osman. Illustrations by Ayman Osman. Published article has been reprinted with the permission of the copyright holder. Printed in Sweden by LiU-Tryck, Linköping, Sweden, 2017 ISBN: 978-91-7685-472-3 ISSN: 0345-0082 To my mother 3 Supervisor: Simin Mohseni, Associate Professor. Department of Clinical and Experimental Medicine (IKE), Linköping University Linköping, Sweden. Co-supervisor: Lars Dahlin, Professor and Senior Consultant in Hand Surgery. Department of Translational Medicine - Hand Surgery, Faculty of Medicine, Lund University. David Engblom, Associate Professor. Department of Clinical and Experimental Medicine (IKE), Linköping University Linköping, Sweden. Maria Turkina, Associate Professor. Department of Clinical and Experimental Medicine (IKE), Linköping University, Linköping, Sweden. Faculty opponent: Kaj Fried, Professor. Department of Neuroscience (Neuro), Karolinska Institute, Stockholm, Sweden. Committee board: Ebo de Muinck, Professor. Department of Medical and Health Sciences (IMH), Linköping University, Linköping, Sweden. Katarina Kågedal, Associate Professor. Department of Clinical and Experimental Medicine (IKE), Linköping University, Linköping, Sweden. Christian Bjerggaard Vægter, Associate Professor. Department of Biomedicine, Aarhus University, Aarhus, Denmark. 4 ABSTRACT Peripheral neuropathy includes a wide range of diseases affecting millions around the world, and many of these diseases have unknown etiology. Peripheral neuropathy in diabetes represents a large proportion of peripheral neuropathies. Nerve damage can also be caused by trauma. -

Traumatic Injury to Peripheral Nerves
AAEM MINIMONOGRAPH 28 ABSTRACT: This article reviews the epidemiology and classification of traumatic peripheral nerve injuries, the effects of these injuries on nerve and muscle, and how electrodiagnosis is used to help classify the injury. Mecha- nisms of recovery are also reviewed. Motor and sensory nerve conduction studies, needle electromyography, and other electrophysiological methods are particularly useful for localizing peripheral nerve injuries, detecting and quantifying the degree of axon loss, and contributing toward treatment de- cisions as well as prognostication. © 2000 American Association of Electrodiagnostic Medicine. Published by John Wiley & Sons, Inc. Muscle Nerve 23: 863–873, 2000 TRAUMATIC INJURY TO PERIPHERAL NERVES LAWRENCE R. ROBINSON, MD Department of Rehabilitation Medicine, University of Washington, Seattle, Washington 98195 USA EPIDEMIOLOGY OF PERIPHERAL NERVE TRAUMA company central nervous system trauma, not only Traumatic injury to peripheral nerves results in con- compounding the disability, but making recognition siderable disability across the world. In peacetime, of the peripheral nerve lesion problematic. Of pa- peripheral nerve injuries commonly result from tients with peripheral nerve injuries, about 60% have 30 trauma due to motor vehicle accidents and less com- a traumatic brain injury. Conversely, of those with monly from penetrating trauma, falls, and industrial traumatic brain injury admitted to rehabilitation accidents. Of all patients admitted to Level I trauma units, 10 to 34% have associated peripheral nerve 7,14,39 centers, it is estimated that roughly 2 to 3% have injuries. It is often easy to miss peripheral nerve peripheral nerve injuries.30,36 If plexus and root in- injuries in the setting of central nervous system juries are also included, the incidence is about 5%.30 trauma. -

Pathophysiology of Peripheral Nerve Injury: a Brief Review
Neurosurg Focus 16 (5):Article 1, 2004, Click here to return to Table of Contents Pathophysiology of peripheral nerve injury: a brief review MARK G. BURNETT, M.D., AND ERIC L. ZAGER, M.D. Department of Neurosurgery, Hospital of the University of Pennsylvania, Philadelphia, Pennsylvania Clinicians caring for patients with brachial plexus and other nerve injuries must possess a clear understanding of the peripheral nervous system’s response to trauma. In this article, the authors briefly review peripheral nerve injury (PNI) types, discuss the common injury classification schemes, and describe the dynamic processes of degeneration and rein- nervation that characterize the PNI response. KEY WORDS • peripheral nerve • nerve injury • neurapraxia • axonotmesis • neurotmesis After a nerve is injured in the periphery, a complex and Lacerations such as those created by a knife blade are finely regulated sequence of events commences to remove another common PNI type, comprising 30% of serious in- the damaged tissue and begin the reparative process. Un- juries in some series.5 Whereas these can be complete like cellular repair in other areas of the body, the response transections, more often some nerve element of continuity of the peripheral nerve to injury does not involve mitosis remains. Because most research models involve a lacer- and cellular proliferation. Our understanding of the peri- ation-type injury mechanism because it is easily repro- pheral nerve regeneration process has increased signifi- duced, the details of nerve degeneration and regeneration cantly during the past several decades concurrent with discussed in this article are perhaps most representative of advances in cellular and molecular biology. -

OIICS Manual 2012
SECTION 4.1 Nature of Injury or Illness Index *-Asterisks denote a summary level code not assigned to individual cases. _____________________________________________________________________________________________ 01/12 447 NATURE CODE INDEX A 2831 Acne 2831 Acne varioliformis 3221 Abacterial meningitis 3211 Acquired immune deficiency syndrome 253 Abdominal hernia from repeated exertions (AIDS)—diagnosed 124 Abdominal hernia from single or short term 3199 Actinomycotic infections exertion 2819 Acute abscess of lymph gland or node 5174 Abdominal pain, unspecified 2359 Acute and subacute endocarditis 521 Abnormal blood-gas level 241 Acute bronchitis and bronchiolitis 521 Abnormal blood-lead level 2341 Acute cor pulmonale 525 Abnormal electrocardiogram (EKG, ECG), 195* Acute dermatitis electroencephalogram (EEG), 2819 Acute lymphadenitis electroretinogram (ERG) 2351 Acute myocarditis 52* Abnormal findings 2359 Acute pericarditis 521 Abnormal findings from examination of 2342 Acute pulmonary artery or vein embolism, blood nontraumatic 522 Abnormal findings from examination of 241 Acute respiratory infections (including urine common cold) 525 Abnormal findings from function studies 2422 Adenoids—chronic condition 526* Abnormal findings from histological and 6212 Adjustment disorder immunological studies 1731 Aero-otitis media 5269 Abnormal findings from histological and 1732 Aero-sinusitis immunological studies, n.e.c. 21 Agranulocytosis and neutropenia 5260 Abnormal findings from histological and 3212 AIDS-like syndrome immunological studies, unspecified 3212 AIDS-related complex (ARC) 523 Abnormal findings from body 3211 AIDS (acquired immune deficiency substances other than blood and urine syndrome)—diagnosed 524 Abnormal findings from radiological and 399 Ainhum other examination of body structure 1733 Air or gas embolisms due to diving 520 Abnormal findings, unspecified 1738 Air pressure effects, multiple 5129 Abnormal gait 1739 Air pressure effects, n.e.c. -

Enhancement of Peripheral Nerve Regeneration with Controlled Release of Glial Cell Line-Derived Neurotrophic Factor (GDNF)
Enhancement of peripheral nerve regeneration with controlled release of glial cell line-derived neurotrophic factor (GDNF) by Kasra Tajdaran B.Sc., University of Toronto, 2013 A thesis submitted in conformity with the requirements for the degree of master of applied science Institute for Biomaterials and Biomedical Engineering University of Toronto © Copyright by Kasra Tajdaran, 2015 Enhancement of peripheral nerve regeneration with controlled release of glial cell line-derived neurotrophic factor (GDNF) Kasra Tajdaran Master of Applied Science (MASc) Institute of Biomaterials and Biomedical Engineering University of Toronto 2015 Abstract Nerve injuries cause severe disability. The present investigational drug delivery strategies for enhancing peripheral nerve regeneration after nerve transection are not yet clinically translatable due to lack of efficiency or biocompatibility. We developed a local delivery system using drug-loaded poly(lactic-co-glycolic acid) (PLGA) microspheres (MS) embedded in a fibrin gel. This drug delivery system (DDS) could be applied at the nerve injury site to deliver exogenous glial cell line-derived neurotrophic factor (GDNF) to the regenerating axons. We used our developed DDS to enhance nerve regeneration in clinically applicable models of severe nerve injuries, including cases with delayed nerve repair and with large nerve defects. ii Declaration of co-authorship The original scientific content of the thesis is comprised of two articles that are submitted to peer-reviewed internationally recognized journals. In both cases these contributions were primarily the work of Kasra Tajdaran. The contributions of the co-authors are declared in the following sections in conformity with the requirements for the degree of Master’s of Applied Science. -

Our Experience on Temporal Bone Fractures: Retrospective Analysis of 141 Cases
Journal of Clinical Medicine Article Our Experience on Temporal Bone Fractures: Retrospective Analysis of 141 Cases Filippo Ricciardiello 1 , Salvatore Mazzone 1,* , Giuseppe Longo 2, Giuseppe Russo 1, Enrico Piccirillo 3, Giuliano Sequino 1, Michele Cavaliere 4, Nunzio Accardo 1, Flavia Oliva 1, Pasquale Salomone 1, Marco Perrella 5, Fabio Zeccolini 6, Domenico Romano 1, Flavia Di Maro 7 , Pasquale Viola 8 , Rosario Cifali 5, Francesco Muto 6 and Jacopo Galli 9 1 Ear Nose Throt Departement AORN Cardarelli, 80100 Napoli, Italy; fi[email protected] (F.R.); [email protected] (G.R.); [email protected] (G.S.); [email protected] (N.A.); fl[email protected] (F.O.); [email protected] (P.S.); [email protected] (D.R.) 2 General Menager AORN Cardarelli, 80100 Napoli, Italy; [email protected] 3 Department of Otology & Skull Base Surgery, Gruppo Otologico, 29121 Piacenza, Italy; [email protected] 4 Unit of Otorhinolaryngology, Department of Neuroscience, Federico II University Hospital, 80138 Napoli, Italy; [email protected] 5 Anesthesiology and Reanimation Department AORN Cardarelli, 80100 Napoli, Italy; [email protected] (M.P.); [email protected] (R.C.) 6 Radiology Department AORN Cardarelli, 80100 Napoli, Italy; [email protected] (F.Z.); [email protected] (F.M.) 7 Otolaryngology-Head and Neck Surgery Department, University Hospital of Verona, 37132 Verona, Italy; fl[email protected] 8 Unit of Audiology, Department of Experimental and Clinical Medicine, Magna Graecia University, 88100 Catanzaro, Italy; [email protected] 9 Institute of Otolaryngology, Head and Neck Surgery, School of Medicine and Surgery, Università Cattolica del Sacro Cuore, 00168 Rome, Italy; [email protected] * Correspondence: [email protected]; Tel.: +39-3338668858 Citation: Ricciardiello, F.; Mazzone, S.; Longo, G.; Russo, G.; Piccirillo, E.; Abstract: Temporal bone fractures are a common lesion of the base of the skull. -

A Classification of Nerve Injuries by H
BRITISH MEDICAL JOURNAL LONDON SATURDAY AUGUST 29 1942 A CLASSIFICATION OF NERVE INJURIES BY H. J. SEDDON, D.M., F.R.C.S. Nu,ffield Professor of Orthopaedic Surgery, University of Oxford In 1863, during the fiercest days of the American Civil War, In contemporary clinical practice there is no question what- Dr. W. A. Hammond, Surgeon-General to the Federal ever that in frequency and importance the first group of Armies, ordered the establishment of a unit for the observa- phenomena far outweigh the second, and it is with loss of tion and treatment of injuiries of the nervous system. A function alone that this classification is concerned. signal result of this instruction was the appearance in 1864 Although, as has been said, there is a fairly clear under- standing among clinicians and experimental workers about the of a little book by Weir Mitchell, Morehouse, and Keen, sequence of events following complete anatomical division of Gunshot Wounds and Other Injuries of Nerves. This a peripheral nerve, there is considerable confusion when we gave an account of the first organized study of nerve come to examine those numerous cases in which function is injuries, and in it we find not only the most perfect descrip- lost although anatomical continuity of the nerve is more or tion of causalgia, a condition that will always remain less preserved. No clear distinction has been drawn between associated with the name of Weir Mitchell, but descrip- those intraneural lesions that are followed by complete tions of nerve injuries as we see them to-day.