Cholesterol Loading Suppresses the Atheroinflammatory Gene
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Genome-Wide Analysis of ATP-Binding
Tian et al. BMC Genomics (2017) 18:330 DOI 10.1186/s12864-017-3706-6 RESEARCH ARTICLE Open Access Genome-wide analysis of ATP-binding cassette (ABC) transporters in the sweetpotato whitefly, Bemisia tabaci Lixia Tian1, Tianxue Song2, Rongjun He1, Yang Zeng1, Wen Xie1, Qingjun Wu1, Shaoli Wang1, Xuguo Zhou3* and Youjun Zhang1* Abstract Background: ABC transporter superfamily is one of the largest and ubiquitous groups of proteins. Because of their role in detoxification, insect ABC transporters have gained more attention in recent years. In this study, we annotated ABC transporters from a newly sequenced sweetpotato whitefly genome. Bemisia tabaci Q biotype is an emerging global invasive species that has caused extensive damages to field crops as well as ornamental plants. Results: A total of 55 ABC transporters containing all eight described subfamilies (A to H) were identified in the B. tabaci Q genome, including 8 ABCAs, 3 ABCBs, 6 ABCCs, 2 ABCDs, 1 ABCE, 3 ABCFs, 23 ABCGs and 9 ABCHs. In comparison to other species, subfamilies G and H in both phloem- and blood-sucking arthropods are expanded. The temporal expression profiles of these 55 ABC transporters throughout B. tabaci developmental stages and their responses to imidacloprid, a neonicotinoid insecticide, were investigated using RNA-seq analysis. Furthermore, the mRNA expression of 24 ABC transporters (44% of the total) representing all eight subfamilies was confirmed by the quantitative real-time PCR (RT-qPCR). Furthermore, mRNA expression levels estimated by RT-qPCR and RNA-seq analyses were significantly correlated (r =0.684,p <0.01). Conclusions: It is the first genome-wide analysis of the entire repertoire of ABC transporters in B. -

Evolutionary and Functional Studies on Microsporidian ATP-Binding
Infection, Genetics and Evolution 68 (2019) 136–144 Contents lists available at ScienceDirect Infection, Genetics and Evolution journal homepage: www.elsevier.com/locate/meegid Evolutionary and functional studies on microsporidian ATP-binding T cassettes: Insights into the adaptation of microsporidia to obligated intracellular parasitism Qiang Hea, Charles R. Vossbrinckc, Qiong Yangd, Xian-Zhi Menga, Jian Luoa, Guo-Qing Pana, ⁎ ⁎ Ze-Yang Zhoua,b, , Tian Lia, a State Key Laboratory of Silkworm Genome Biology, Southwest University, Chongqing 400715, China b College of Life Science, Chongqing Normal University, Chongqing 400047, China c Department of Soil and Water, The Connecticut Agricultural Experiment Station, 123 Huntington Street, New Haven, CT 06511, USA d Sericulture and Agri-Food Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, China ARTICLE INFO ABSTRACT Keywords: ATP-binding cassette (ABC) transporters comprise the largest family of transmembrane proteins and are found in Microsporidian all domains of life. The ABCs are involved in a variety of biological processes and as exporters play important ABC transporter roles in multidrug resistance. However, the ABC transporters have not been addressed in microsporidia, which Genome are a very large group of obligate intracellular parasites that can infect nearly all animals, including humans. Evolution Here, a total of 234 ABC transporters were identified from 18 microsporidian genomes and classified intofive Nosema bombycis subfamilies, including 74 ABCBs, 2 ABCCs, 18 ABCEs, 15 ABCFs, 102 ABCGs and 23 uncategorized members. Two subfamilies, ABCA and ABCD, are found in most organisms, but lost in microsporidia. Phylogenetic analysis indicated that microsporidian ABCB and ABCG subfamilies expanded by recent gene duplications, which re- sulted in the two largest subfamilies in microsporidia. -

Polymorphisms of the Multidrug Pump ABCG2: a Systematic Review of Their Effect on Protein Expression, Function, and Drug Pharmacokinetics
1521-009X/46/12/1886–1899$35.00 https://doi.org/10.1124/dmd.118.083030 DRUG METABOLISM AND DISPOSITION Drug Metab Dispos 46:1886–1899, December 2018 Copyright ª 2018 by The American Society for Pharmacology and Experimental Therapeutics Minireview Polymorphisms of the Multidrug Pump ABCG2: A Systematic Review of Their Effect on Protein Expression, Function, and Drug Pharmacokinetics Niall Heyes, Parth Kapoor, and Ian D. Kerr School of Life Sciences, Queen’s Medical Centre, University of Nottingham, Nottingham, United Kingdom Received June 12, 2018; accepted September 20, 2018 Downloaded from ABSTRACT The widespread expression and polyspecificity of the multidrug to improve outcomes in cancer patients treated with tyrosine ABCG2 efflux transporter make it an important determinant of the kinase inhibitors, but the reasons for this are yet to be estab- pharmacokinetics of a variety of substrate drugs. Null ABCG2 lished, and this residue’s role in the mechanism of the protein is expression has been linked to the Junior blood group. Polymor- unexplored by current biochemical and structural approaches. phisms affecting the expression or function of ABCG2 may have Research into the less-common polymorphisms is confined to dmd.aspetjournals.org clinically important roles in drug disposition and efficacy. The in vitro studies, with several polymorphisms shown to decrease most well-studied single nucleotide polymorphism (SNP), Q141K resistance to anticancer agents such as SN-38 and mitoxantrone. (421C>A), is shown to decrease ABCG2 expression and activity, In this review, we present a systematic analysis of the effects of resulting in increased total drug exposure and decreased re- ABCG2 polymorphisms on ABCG2 function and drug pharmaco- sistance to various substrates. -

Identification of Candidate ATP-Binding Cassette Transporter
www.nature.com/scientificreports OPEN Identifcation of candidate ATP- binding cassette transporter gene family members in Diaphorina citri (Hemiptera: Psyllidae) via adult tissues transcriptome analysis Zhengbing Wang 1, Fajun Tian1, Lijun Cai2, Jie Zhang2, Jiali Liu1 & Xinnian Zeng1* The ATP-binding cassette (ABC) transporters exist in all living organisms and play major roles in various biological functions by transporting a wide variety of substrates across membranes. The functions of ABC transporters in drug resistance have been extensively studied in vertebrates; however, they are rarely characterized in agricultural pests. The Asian citrus psyllid, Diaphorina citri, is one of the most damaging pests of the Citrus genus because of its transmission of Huanglongbing, also known as Yellow Dragon disease. In this study, the next-generation sequencing technique was applied to research the ABC transporters of D. citri. Fifty-three ABC transporter genes were found in the RNA-Seq data, and among these ABC transporters, 4, 4, 5, 2, 1, 4, 18 and 15 ABC proteins belonged to the ABCA-ABCH subfamilies, respectively. Diferent expression profles of 52 genes between imidacloprid-resistant and imidacloprid-susceptible strains were studied by qRT-PCR; 5 ABCGs and 4 ABCHs were signifcantly upregulated in the imidacloprid-resistant strain. In addition, fve of the nine upregulated genes were widely expressed in adult tissues in spatial expression analysis. The results suggest that these genes may play key roles in this phenotype. In general, this study contributed to our current understanding of D. citri resistance to insecticides. Te ATP-binding cassette (ABC) transporter family is one of the largest families of membrane proteins and uni- versally exists in all living organisms on Earth1. -

ABCG4: a Novel Human White Family ABC-Transporter Expressed in the Brain and Eye
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Biochimica et Biophysica Acta 1591 (2002) 175–179 www.bba-direct.com Short sequence-paper ABCG4: a novel human white family ABC-transporter expressed in the brain and eye Susan Oldfield *, Christopher A. Lowry, Jon Ruddick, Stafford L. Lightman University Research Centre for Neuroendocrinology, BRI, University of Bristol, Marlborough Street, Bristol BS2 8HW, UK Received 24 July 2001; received in revised form 11 April 2002; accepted 24 May 2002 Abstract White family transporters are a group of ATP-binding cassette (ABC) proteins that show sequence homology to the Drosophila white gene product. The Drosophila protein is a subunit of heterodimeric transporters of precursors for eye-pigment synthesis. A novel, human member of this family (ABCG4) has been identified. Northern blotting shows that ABCG4 is expressed specifically in the brain and the eye. D 2002 Elsevier Science B.V. All rights reserved. Keywords: White gene; ABC-transporter; ABCG1; Sterol ATP-binding cassette (ABC) transporters form a large and its murine counterpart is known as ABCG1 or ABC8. superfamily of proteins that mediate the transport of a large Northern blotting has shown that the mRNAs encoding variety of diverse molecules across biological membranes in ABCG1 are present in several tissues, predominantly brain, an ATP-dependent manner [1]. The functional transporters spleen and lung [10]. contain two ATP-binding folds and two transmembrane The function of ABCG1 is unknown, although there is domains each with, usually, five or six membrane spanning evidence that suggests that it may have a role in cholesterol helices. -

Transcriptional and Post-Transcriptional Regulation of ATP-Binding Cassette Transporter Expression
Transcriptional and Post-transcriptional Regulation of ATP-binding Cassette Transporter Expression by Aparna Chhibber DISSERTATION Submitted in partial satisfaction of the requirements for the degree of DOCTOR OF PHILOSOPHY in Pharmaceutical Sciences and Pbarmacogenomies in the Copyright 2014 by Aparna Chhibber ii Acknowledgements First and foremost, I would like to thank my advisor, Dr. Deanna Kroetz. More than just a research advisor, Deanna has clearly made it a priority to guide her students to become better scientists, and I am grateful for the countless hours she has spent editing papers, developing presentations, discussing research, and so much more. I would not have made it this far without her support and guidance. My thesis committee has provided valuable advice through the years. Dr. Nadav Ahituv in particular has been a source of support from my first year in the graduate program as my academic advisor, qualifying exam committee chair, and finally thesis committee member. Dr. Kathy Giacomini graciously stepped in as a member of my thesis committee in my 3rd year, and Dr. Steven Brenner provided valuable input as thesis committee member in my 2nd year. My labmates over the past five years have been incredible colleagues and friends. Dr. Svetlana Markova first welcomed me into the lab and taught me numerous laboratory techniques, and has always been willing to act as a sounding board. Michael Martin has been my partner-in-crime in the lab from the beginning, and has made my days in lab fly by. Dr. Yingmei Lui has made the lab run smoothly, and has always been willing to jump in to help me at a moment’s notice. -
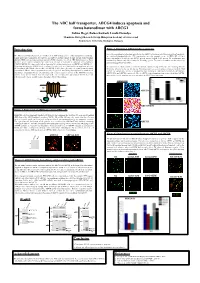
The ABC Half Transporter, ABCG4 Induces Apoptosis and Forms Heterodimer with ABCG1
The ABC half transporter, ABCG4 induces apoptosis and forms heterodimer with ABCG1 Zoltán Hegyi, Balázs Sarkadi, László Homolya Membrane Biology Research Group, Hungarian Academy of Sciences and Semmelweis University, Budapest, Hungary Introduction Figure 3. Functional ABCG4 induces apoptosis Since the morphological alterations observed in the ABCG4-expressing cells were indicative of apoptosis, The ABCG1 and ABCG4 proteins are members of the ATP binding cassette (ABC) transporter G subfamily. we examined phosphatidyl-serine (PS) translocation, an early apoptotic event in HEK293H cell lines Unlike most ABC transporters, the ABCG1 and ABCG4 proteins consist of only one nucleotide binding transiently transfected with the wt ABCG4 and its inactive mutant (KM) variant. PS translocation was domain (NBD) and one transmembrane domain (TMD), therefore are called ABC half-transporters. Some monitored by fluorescently labeled Annexin V binding (green). The total cell number was determined by members of the ABCG subfamily have proven to function as homodimers (ABCG2) or heterodimers nuclear staining by Hoechst (blue). (ABCG5/ABCG8). Previous results indicated potential heterodimerization between ABCG1 and ABCG4 (1). Regarding their function, ABCG1 has been proposed to play a role in cellular lipid/sterol regulation, whereas We found that the cultures transfected with wt ABCG4 contained a large number of cells exhibiting Annexin the function of ABCG4, the closest relative of ABCG1, is still elusive. Recently, we reported that functional V binding, whereas hardly any labeling for PS translocation was seen in cultures transfected with the KM expression of ABCG1 induces apoptosis in several cell types (2). Our finding was supported by rounded cell mutant. For comparison, Annexin V binding was also examined in cells transfected with the wt ABCG1, morphology, phosphatidyl-serine externalization, and elevated caspase 3 activity in the ABCG1-expressing ABCG1-KM, and ABCG2, respectively. -

Influence of ABCA1 and ABCA7 on the Lipid Microenvironment of the Plasma Membrane
I Influence of ABCA1 and ABCA7 on the lipid microenvironment of the plasma membrane Dissertation zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) im Fach Biologie eingereicht an der Mathematisch-Naturwissenschaftlichen Fakultät I der Humboldt-Universität zu Berlin von Dottore Magistrale in Biotecnologie Industriali Anna Pia Plazzo geb. 17.10.1981 in San Giovanni Rotondo, Italien Präsident der Humboldt-Universität zu Berlin Prof. Dr. Dr. h.c. Christoph Markschies Dekan der Mathematisch-Naturwissenschaftlichen Fakultät I Prof. Dr. Lutz-Helmut Schön Gutachter: 1. Prof. Dr. Andreas Herrmann 2. Prof. Dr. Thomas Günther-Pomorski 3. Prof. Dr. Thomas Eitinger Tag der mündlichen Prüfung: 12.06.2009 II Alla mia famiglia III Zusammenfassung Der ABC-Transporter ABCA1 ist unmittelbar in die zelluläre Lipidhomeostasie einbezogen, in dem er die Freisetzung von Cholesterol an plasmatische Rezeptoren, wie ApoA-I, vermittelt. Trotz intensiver Untersuchungen ist dieser molekulare Mechanismus nicht verstanden. Verschiedene Studien deuten daraufhin, dass durch die Aktivität von ABCA1 bedingte Veränderungen in der Lipidphase der äußeren Hälfte der Plasmamembran (PM) wichtig für die Freisetzung des Cholesterols sind. In der vorliegenden Arbeit wird die Lipidumgebung von ABCA1 in der PM lebender Säugetierzellen unter Anwendung der Fluoreszenzlebenszeitmikroskopie von fluoreszierenden Lipidsonden untersucht. Es wurde eine breite Verteilung der Fluoreszenzlebenszeiten der Sonden gefunden, die sensitiv gegenüber Veränderungen der lateralen und transversalen Organisation der Lipide ist. Im Einklang mit Studien an riesengroßen unilamellaren Vesikeln und Plasmamembranvesikeln weisen unsere Ergebnisse die Existenz einer größeren Vielfalt submikroskopischer Lipiddomänen auf. Die FLIM-Untersuchungen an ABCA1 exprimierenden HeLa-Zellen weisen eine die Lipidphase destabilisierende Funktion des Transportes aus. Dieses wurde unterstützt durch die Lipidanalyse von Fraktionen der PM. -

Role in Restricting the Brain Entry of Amyloid-Β Peptide
www.nature.com/scientificreports OPEN Expression and function of Abcg4 in the mouse blood-brain barrier: role in restricting the brain entry of Received: 27 March 2017 Accepted: 27 September 2017 amyloid-β peptide Published: xx xx xxxx Agnès Dodacki1,2,3, Matthew Wortman4, Bruno Saubaméa1,2,3, Stéphanie Chasseigneaux1,2,3, Sophie Nicolic1,2,3, Nathalie Prince1,2,3, Murielle Lochus1,2,3, Anne-Laure Raveu1,2,3, Xavier Declèves1,2,3, Jean-Michel Scherrmann1,2,3, Shailendra B. Patel 4 & Fanchon Bourasset1,2,3 ABCG4 is an ATP-binding cassette transmembrane protein which has been shown, in vitro, to participate in the cellular efux of desmosterol and amyloid-β peptide (Aβ). ABCG4 is highly expressed in the brain, but its localization and function at the blood-brain barrier (BBB) level remain unknown. We demonstrate by qRT-PCR and confocal imaging that mouse Abcg4 is expressed in the brain capillary endothelial cells. Modelling studies of the Abcg4 dimer suggested that desmosterol showed thermodynamically favorable binding at the putative sterol-binding site, and this was greater than for cholesterol. Additionally, unbiased docking also showed Aβ binding at this site. Using a novel Abcg4-defcient mouse model, we show that Abcg4 was able to export Aβ and desmosterol at the BBB level and these processes could be inhibited by probucol and L-thyroxine. Our assay also showed that desmosterol antagonized the export of Aβ, presumably as both bind at the sterol-binding site on Abcg4. We show for the frst time that Abcg4 may function in vivo to export Aβ at the BBB, in a process that can be antagonized by its putative natural ligand, desmosterol (and possibly cholesterol). -

Connecting Cholesterol Efflux Factors to Lung Cancer Biology And
International Journal of Molecular Sciences Review Connecting Cholesterol Efflux Factors to Lung Cancer Biology and Therapeutics Maria Maslyanko †, Ryan D. Harris † and David Mu * Leroy T. Canoles Jr. Cancer Research Center, Department of Microbiology and Molecular Cell Biology, Eastern Virginia Medical School, Norfolk, VA 23501, USA; [email protected] (M.M.); [email protected] (R.D.H.) * Correspondence: [email protected] † Equal contribution. Abstract: Cholesterol is a foundational molecule of biology. There is a long-standing interest in understanding how cholesterol metabolism is intertwined with cancer biology. In this review, we focus on the known connections between lung cancer and molecules mediating cholesterol efflux. A major take-home lesson is that the roles of many cholesterol efflux factors remain underexplored. It is our hope that this article would motivate others to investigate how cholesterol efflux factors contribute to lung cancer biology. Keywords: lung cancer; cholesterol efflux; ABCA1; ABCG1; Apo AI; miRNA; miR-33a; miR-200b-3p; LRPs; LAL; NPC1; STARD3; SMPD1; NCEH1; SR-BI; TTF-1; drug resistance; cisplatin 1. Introduction Citation: Maslyanko, M.; Harris, Cholesterol is essential for cell viability and cell membrane integrity. Cholesterol is R.D.; Mu, D. Connecting Cholesterol also a precursor to many physiologically important hormones. The interest in the intercon- Efflux Factors to Lung Cancer Biology nection between cholesterol metabolism and cancer is best illustrated by the abundance of and Therapeutics. Int. J. Mol. Sci. literature on this subject matter. A quick search of PubMed.gov (accessed on 4 June 2021) 2021, 22, 7209. https://doi.org/ using the two key words (Cholesterol AND Cancer) retrieves over 20,000 publications. -

High ABCG4 Expression Is Associated with Poor Prognosis in Non-Small-Cell Lung Cancer Patients Treated with Cisplatin-Based Chemotherapy
RESEARCH ARTICLE High ABCG4 Expression Is Associated with Poor Prognosis in Non-Small-Cell Lung Cancer Patients Treated with Cisplatin-Based Chemotherapy Guang Yang☯, Xue-Jiao Wang☯, Li-Jun Huang☯, Yong-An Zhou, Feng Tian, Jin-Bo Zhao, Peng Chen, Bo-Ya Liu, Miao-Miao Wen, Xiao-Fei Li*, Zhi-Pei Zhang* Department of Thoracic Surgery, Tangdu Hospital, the Fourth Military Medical University, Xi’an, 710038, China ☯ These authors contributed equally to this work. * [email protected] (XFL); [email protected] (ZPZ) Abstract ATP-binding cassette (ABC) transporters are associated with poor response to chemother- OPEN ACCESS apy, and confer a poor prognosis in various malignancies. However, the association Citation: Yang G, Wang X-J, Huang L-J, Zhou Y-A, between the expression of the ABC sub-family G member 4 (ABCG4) and prognosis in Tian F, Zhao J-B, et al. (2015) High ABCG4 patients with non-small-cell lung cancer (NSCLC) remains unclear. NSCLC tissue samples Expression Is Associated with Poor Prognosis in Non-Small-Cell Lung Cancer Patients Treated with (n = 140) and normal lung tissue samples (n = 90) were resected from patients with stage II Cisplatin-Based Chemotherapy. PLoS ONE 10(8): to IV NSCLC between May 2004 and May 2009. ABCG4 mRNA and protein expressions e0135576. doi:10.1371/journal.pone.0135576 were detected by RT-PCR, western blot, and immunohistochemistry. Patients received four st Editor: Gergely Szakacs, Hungarian Academy of cycles of cisplatin-based post-surgery chemotherapy and were followed up until May 31 , Sciences, HUNGARY 2014. ABCG4 positivity rate was higher in NSCLC than in normal lung tissues (48.6% vs. -

Frequent Down-Regulation of ABC Transporter Genes in Prostate Cancer
Demidenko et al. BMC Cancer (2015) 15:683 DOI 10.1186/s12885-015-1689-8 RESEARCH ARTICLE Open Access Frequent down-regulation of ABC transporter genes in prostate cancer Rita Demidenko1, Deividas Razanauskas1, Kristina Daniunaite1, Juozas Rimantas Lazutka1, Feliksas Jankevicius2,3 and Sonata Jarmalaite1* Abstract Background: ATP-binding cassette (ABC) transporters are transmembrane proteins responsible for the efflux of a wide variety of substrates, including steroid metabolites, through the cellular membranes. For better characterization of the role of ABC transporters in prostate cancer (PCa) development, the profile of ABC transporter gene expression was analyzed in PCa and noncancerous prostate tissues (NPT). Methods: TaqMan Low Density Array (TLDA) human ABC transporter plates were used for the gene expression profiling in 10 PCa and 6 NPT specimens. ABCB1 transcript level was evaluated in a larger set of PCa cases (N = 78) and NPT (N = 15) by real-time PCR, the same PCa cases were assessed for the gene promoter hypermethylation by methylation-specific PCR. Results: Expression of eight ABC transporter genes (ABCA8, ABCB1, ABCC6, ABCC9, ABCC10, ABCD2, ABCG2, and ABCG4) was significantly down-regulated in PCa as compared to NPT, and only two genes (ABCC4 and ABCG1) were up-regulated. Down-regulation of ABC transporter genes was prevalent in the TMPRSS2-ERG-negative cases. A detailed analysis of ABCB1 expression confirmed TLDA results: a reduced level of the transcript was identified in PCa in comparison to NPT (p = 0.048). Moreover, the TMPRSS2-ERG-negative PCa cases showed significantly lower expression of ABCB1 in comparison to NPT (p = 0.003) or the fusion-positive tumors (p = 0.002).