Ecoloyy Coregonus Lavaretus
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Coregonus Lavaretus (L.)) Caused by Competitor Invasion
Speciation Reversal in European Whitefish (Coregonus lavaretus (L.)) Caused by Competitor Invasion Shripathi Bhat1*, Per-Arne Amundsen1, Rune Knudsen1, Karl Øystein Gjelland3, Svein-Erik Fevolden1, Louis Bernatchez2, Kim Præbel1 1 Department of Arctic and Marine Biology, University of Tromsø, Tromsø, Norway, 2 Institut de Biologie Inte´grative et des Syste`mes (IBIS), Universite´ Laval, Que´bec, Canada, 3 Norwegian Institute for Nature Research, Tromsø, Norway Abstract Invasion of exotic species has caused the loss of biodiversity and imparts evolutionary and ecological changes in the introduced systems. In northern Fennoscandia, European whitefish (Coregonus lavaretus (L.)) is a highly polymorphic species displaying adaptive radiations into partially reproductively isolated and thus genetically differentiated sympatric morphs utilizing the planktivorous and benthivorous food niche in many lakes. In 1993, Lake Skrukkebukta was invaded by vendace (Coregonus albula (L.)) which is a zooplanktivorous specialist. The vendace displaced the densely rakered whitefish from its preferred pelagic niche to the benthic habitat harbouring the large sparsely rakered whitefish. In this study, we investigate the potential influence of the vendace invasion on the breakdown of reproductive isolation between the two whitefish morphs. We inferred the genotypic and phenotypic differentiation between the two morphs collected at the arrival (1993) and 15 years after (2008) the vendace invasion using 16 microsatellite loci and gill raker numbers, the most distinctive adaptive phenotypic trait between them. The comparison of gill raker number distributions revealed two modes growing closer over 15 years following the invasion. Bayesian analyses of genotypes revealed that the two genetically distinct whitefish morphs that existed in 1993 had collapsed into a single population in 2008. -

First Record of a Coregonid Fish Species, Coregenus Albula (Linnaeus, 1758) (Salmoniformes: Salmonidae) in Aktaş Lake Shared Between Turkey and Georgia
J. Black Sea/Mediterranean Environment Vol. 25, No. 3: 325-332 (2019) SHORT COMMUNICATION First record of a coregonid fish species, Coregenus albula (Linnaeus, 1758) (Salmoniformes: Salmonidae) in Aktaş Lake shared between Turkey and Georgia Sedat V. Yerli Department of Biology, Hacettepe University, SAL, Beytepe, Ankara, TURKEY Corresponding author: [email protected] Abstract The genus Coregenus (Salmoniformes: Salmonidae) was recently considered not to be represented in Turkey. European cisco or vendace, Coregonus albula (Linnaeus, 1758) was reported for the first time for Turkey in this article with fifteen samples in Aktaş Lake, Ardahan. This species should be added to the checklist of Turkish fish fauna. Turkish name is proposed as “Akbalık” for this species. Keywords: Coregonus albula, first record, Aktaş Lake, Kartsakhi, alkaline lake, Georgia, Turkey Received: 30.10.2019, Accepted: 26.11.2019 Vendace or European cisco Coregonus albula (Linnaeus, 1758) is a native species for northern Europe. Berg (1948) reported the distribution of this species its morphological measurements in the former USSR and adjacent countries. Froese and Pauly (2019) summarized the natural distribution of vendace as Baltic basin, several lakes of upper Volga drainage; some lakes of White Sea basin and North Sea basin east of Elbe drainage; anadromous in Gulf of Finland and marine in northernmost freshened part of Gulf of Bothnia between Finland and Sweden; in Lake Inari, northern Finland; lower Rhine (now extirpated). The vendace was introduced, intentionally in some countries in Europe and United States of America. Vendace was introduced in 1959, 1982-1987 in the Irtysh River Basin and in 1960-61 in Lake Balkhash in Kazakhstan (Mitrofanov and Petr 1999). -

Coregonus Lavaretus Complex 1.4 Alternative Species Scientific Name 1.5 Common Name (In National Language) Whitefish 2
European Community Directive on the Conservation of Natural Habitats and of Wild Fauna and Flora (92/43/EEC) Fourth Report by the United Kingdom under Article 17 on the implementation of the Directive from January 2013 to December 2018 Supporting documentation for the conservation status assessment for the species: S6353 ‐ WhitefishCoregonus ( lavaretus) SCOTLAND IMPORTANT NOTE ‐ PLEASE READ • The information in this document is a country‐level contribution to the UK Reporton the conservation status of this species, submitted to the European Commission aspart of the 2019 UK Reporting under Article 17 of the EU Habitats Directive. • The 2019 Article 17 UK Approach document provides details on how this supporting information was used to produce the UK Report. • The UK Report on the conservation status of this species is provided in a separate doc‐ ument. • The reporting fields and options used are aligned to those set out in the European Com‐ mission guidance. • Explanatory notes (where provided) by the country are included at the end. These pro‐ vide an audit trail of relevant supporting information. • Some of the reporting fields have been left blank because either: (i) there was insuffi‐ cient information to complete the field; (ii) completion of the field was not obligatory; (iii) the field was not relevant to this species (section 12 Natura 2000 coverage forAnnex II species) and/or (iv) the field was only relevant at UK‐level (sections 9 Future prospects and 10 Conclusions). • For technical reasons, the country‐level future trends for Range, Population and Habitat for the species are only available in a separate spreadsheet that contains all the country‐ level supporting information. -

Lake Superior Food Web MENT of C
ATMOSPH ND ER A I C C I A N D A M E I C N O I S L T A R N A T O I I O T N A N U E .S C .D R E E PA M RT OM Lake Superior Food Web MENT OF C Sea Lamprey Walleye Burbot Lake Trout Chinook Salmon Brook Trout Rainbow Trout Lake Whitefish Bloater Yellow Perch Lake herring Rainbow Smelt Deepwater Sculpin Kiyi Ruffe Lake Sturgeon Mayfly nymphs Opossum Shrimp Raptorial waterflea Mollusks Amphipods Invasive waterflea Chironomids Zebra/Quagga mussels Native waterflea Calanoids Cyclopoids Diatoms Green algae Blue-green algae Flagellates Rotifers Foodweb based on “Impact of exotic invertebrate invaders on food web structure and function in the Great Lakes: NOAA, Great Lakes Environmental Research Laboratory, 4840 S. State Road, Ann Arbor, MI A network analysis approach” by Mason, Krause, and Ulanowicz, 2002 - Modifications for Lake Superior, 2009. 734-741-2235 - www.glerl.noaa.gov Lake Superior Food Web Sea Lamprey Macroinvertebrates Sea lamprey (Petromyzon marinus). An aggressive, non-native parasite that Chironomids/Oligochaetes. Larval insects and worms that live on the lake fastens onto its prey and rasps out a hole with its rough tongue. bottom. Feed on detritus. Species present are a good indicator of water quality. Piscivores (Fish Eaters) Amphipods (Diporeia). The most common species of amphipod found in fish diets that began declining in the late 1990’s. Chinook salmon (Oncorhynchus tshawytscha). Pacific salmon species stocked as a trophy fish and to control alewife. Opossum shrimp (Mysis relicta). An omnivore that feeds on algae and small cladocerans. -

Coregonus Maraena) Ecological Risk Screening Summary
Maraena Whitefish (Coregonus maraena) Ecological Risk Screening Summary U.S. Fish and Wildlife Service, April 2011 Revised, September 2014 and June 2017 Web Version, 09/14/2017 Image: E. Östman. Public domain. Available: http://eol.org/data_objects/26779416. (June 2017). 1 Native Range, and Status in the United States Native Range From Froese and Pauly (2017): “Europe: In the Baltic Sea: Swedish coast (including Bothnian Gulf, not in Gotland); in southern Baltic, extending from the Schlei to Gulf of Finland. Southeast North Sea Basin: Ems, Weser and Elbe drainages 1 and small rivers of Schleswig-Holstein and Denmark. Landlocked in several lakes in Poland, Sweden, and Russia.” Status in the United States From Neilson (2017): “Failed introduction.” “A shipment of 409 individuals from Lake Miedwie (formerly Madue Lake), Poland was stocked in Garnder Lake, Michigan in 1877 (Baird 1879; Todd 1983).” Means of Introductions in the United States From Neilson (2017): “Coregonus maraena, along with other species of Coregonus, was intentionally stocked as a food fish by the U.S. Fish Commission (Todd 1983). According to Baird (1879), 1,000 eggs of C. maraena were shipped from Poland to Michigan in 1877 and hatched in captivity at the State Hatching House in Detroit. A total of 409 of the young fish were stocked in Gardner Lake (Baird 1879; Todd 1983). Baird (1879) considered the stocking an experimental introduction of a European food fish.” Remarks From Neilson (2017): “There is much confusion regarding the identity of whitefish imported from Germany in the late 1800s by the U.S. Fish Commission, primarily due to the uncertain taxonomy and systematics of Coregonus (Kottelat and Freyhof 2007). -

Identification and Modelling of a Representative Vulnerable Fish Species for Pesticide Risk Assessment in Europe
Identification and Modelling of a Representative Vulnerable Fish Species for Pesticide Risk Assessment in Europe Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der RWTH Aachen University zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften genehmigte Dissertation vorgelegt von Lara Ibrahim, M.Sc. aus Mazeraat Assaf, Libanon Berichter: Universitätsprofessor Dr. Andreas Schäffer Prof. Dr. Christoph Schäfers Tag der mündlichen Prüfung: 30. Juli 2015 Diese Dissertation ist auf den Internetseiten der Universitätsbibliothek online verfügbar Erklärung Ich versichere, dass ich diese Doktorarbeit selbständig und nur unter Verwendung der angegebenen Hilfsmittel angefertigt habe. Weiterhin versichere ich, die aus benutzten Quellen wörtlich oder inhaltlich entnommenen Stellen als solche kenntlich gemacht zu haben. Lara Ibrahim Aachen, am 18 März 2015 Zusammenfassung Die Zulassung von Pflanzenschutzmitteln in der Europäischen Gemeinschaft verlangt unter anderem eine Abschätzung des Risikos für Organismen in der Umwelt, die nicht Ziel der Anwendung sind. Unvertretbare Auswirkungen auf den Naturhalt sollen vermieden werden. Die ökologische Risikoanalyse stellt die dafür benötigten Informationen durch eine Abschätzung der Exposition der Organismen und der sich daraus ergebenden Effekte bereit. Die Effektabschätzung beruht dabei hauptsächlich auf standardisierten ökotoxikologischen Tests im Labor mit wenigen, oft nicht einheimischen Stellvertreterarten. In diesen Tests werden z. B. Effekte auf das Überleben, das Wachstum und/oder die Reproduktion von Fischen bei verschiedenen Konzentrationen der Testsubstanz gemessen und Endpunkte wie die LC50 (Lethal Concentrations for 50%) oder eine NOEC (No Observed Effect Concentration, z. B. für Wachstum oder Reproduktionsparameter) abgeleitet. Für Fische und Wirbeltiere im Allgemeinen beziehen sich die spezifischen Schutzziele auf das Überleben von Individuen und die Abundanz und Biomasse von Populationen. -
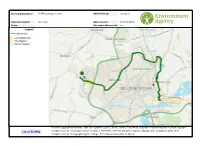
GIS Screening
Screening Results for: IR EPR discharges to water Date Produced: 16/09/2020 Reference number: KP3236YH NGR of search: SP 86093 68828 Notes: Site centre distance (m): 80 https://easimap2. prodds. ntnl/Html5Viewer/I ndex.html? viewer=Permitting &Extent=484159. 913849983, 266371. 936523897, © Crown Copyright and database right 2020. Ordnance Survey licence number 100024198. Copyright © Natural England. © Crown Copyright. 492626. Launch597450017, EasiMap All rights reserved. Countryside Council for Wales, 100018813, 2020. © Environment Agency copyright and / or database rights 2020. 270889. All rights reserved. © Copyright English Heritage. © The National Assembly for Wales. Easimap Screening Results Click here to download Permit Screening distances Screening Results for: IR EPR discharges to water Date produced: 16/09/2020 Reference number: KP3236YH NGR of search: SP 86093 68828 Notes: Site centre distance (m): 80 Data Details Within Search Direction Action Special Protection Areas - England Site Name: Upper Nene Valley Gravel Pits 9813m Downstream in the HRA Stage 1 Form (App11) consultation with NE may be required (SPA) nearest watercourse Ramsar Sites - England Site Name: Upper Nene Valley Gravel Pits 9813m Downstream in the HRA Stage 1 Form (App11) consultation with NE may be required (Ramsar) nearest watercourse Local Wildlife Sites Name: Park Farm Industrial Estate 424m Downstream in the Take account of the requirements of this site. If there is no data available nearest watercourse consult Area EPR Installation Team EPR Team: Installations -

Alaska Arctic Marine Fish Ecology Catalog
Prepared in cooperation with Bureau of Ocean Energy Management, Environmental Studies Program (OCS Study, BOEM 2016-048) Alaska Arctic Marine Fish Ecology Catalog Scientific Investigations Report 2016–5038 U.S. Department of the Interior U.S. Geological Survey Cover: Photographs of various fish studied for this report. Background photograph shows Arctic icebergs and ice floes. Photograph from iStock™, dated March 23, 2011. Alaska Arctic Marine Fish Ecology Catalog By Lyman K. Thorsteinson and Milton S. Love, editors Prepared in cooperation with Bureau of Ocean Energy Management, Environmental Studies Program (OCS Study, BOEM 2016-048) Scientific Investigations Report 2016–5038 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior SALLY JEWELL, Secretary U.S. Geological Survey Suzette M. Kimball, Director U.S. Geological Survey, Reston, Virginia: 2016 For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment—visit http://www.usgs.gov or call 1–888–ASK–USGS. For an overview of USGS information products, including maps, imagery, and publications, visit http://store.usgs.gov. Disclaimer: This Scientific Investigations Report has been technically reviewed and approved for publication by the Bureau of Ocean Energy Management. The information is provided on the condition that neither the U.S. Geological Survey nor the U.S. Government may be held liable for any damages resulting from the authorized or unauthorized use of this information. The views and conclusions contained in this document are those of the authors and should not be interpreted as representing the opinions or policies of the U.S. -

Age, Growth, and Size of Lake Superior Pygmy Whitefish (Prosopium Coulterii) Author(S): Taylor R
Age, Growth, and Size of Lake Superior Pygmy Whitefish (Prosopium coulterii) Author(s): Taylor R. Stewart and Derek H. OgleOwen T. Gorman and Mark R. Vinson Source: The American Midland Naturalist, 175(1):24-36. Published By: University of Notre Dame DOI: http://dx.doi.org/10.1674/amid-175-01-24-36.1 URL: http://www.bioone.org/doi/full/10.1674/amid-175-01-24-36.1 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Am. Midl. Nat. (2016) 175:24–36 Age, Growth, and Size of Lake Superior Pygmy Whitefish (Prosopium coulterii) 1 TAYLOR R. STEWART AND DEREK H. OGLE Department of Natural Resources, Northland College, Ashland, Wisconsin 54806 AND OWEN T. GORMAN AND MARK R. VINSON U. S. Geological Survey, Great Lakes Science Center, Lake Superior Biological Station, Ashland, Wisconsin 54806 ABSTRACT.—Pygmy Whitefish (Prosopium coulterii) are a small, glacial relict species with a disjunct distribution in North America and Siberia. -

Stormy Fish Sex Ian J. Winfield & J. Ben James
Stormy fish sex Ian J. Winfield & J. Ben James Structure of presentation • Introduction • Methods • Results • Summary Introduction Introduction • Spawning success has stringent environmental requirements • Fish are highly selective in their use of spawning habitat in lakes and their tributaries • Lithophilic species require clean gravel or stones onto or amongst which to place their eggs Introduction • Arctic charr (Salvelinus alpinus ) • Schelly (Coregonus lavaretus ) • Vendace (Coregonus albula ) • All three species are lithophilic Introduction Frost (1965) Introduction Frost (1965) Introduction Bolgan et al. (2017) Introduction Introduction Arctic charr (Salvelinus alpinus ) Schelly (Coregonus lavaretus ) Red Tarn Vendace (Coregonus albula ) Introduction • In early December 2015, Storm Desmond delivered extremely high rainfall and subsequent flooding across Cumbria • Extensive and catastrophic impacts in and around many water bodies • Major movements of gravel and other substrates Introduction Introduction Introduction • The objectives of this project were: • To review pre-2015 descriptions of rare fish spawning habitat in Cumbrian lakes • To undertake new post-Storm Desmond surveys of known, putative or potential rare fish spawning habitat at selected lakes • To consider appropriate mitigation measures for sediment control in upstream catchments Methods Methods 14 lakes 9 lakes Red Tarn Surveys between January and March 2017 Methods • Underwater still photography of inshore substrate at depth of 0.5 m • Underwater videography of substrate along inshore to offshore transects at depths between 0.25 m and 10 m Methods • BioBase/EcoSound (www.cibiobase.com) combines consumer hardware and cloud computing • Produces geo-referenced data for bathymetry, macrophytes and bottom hardness • 5 or 6 inshore areas (each c. 100 m by c. 100 m) surveyed at each lake Methods Winfield et al. -

Population Dynamics of Whitefish ( Coregonus Suidteri Fatio) in Artificially Oxygenated Lake Hallwil, with Special Emphasis on L
Diss. ETH No. 13706 Population dynamics of whitefish ( Coregonus suidteri Fatio) in artificially oxygenated Lake Hallwil, with special emphasis on larval mortality and sustainable management Dissertation submitted to the SWISS FEDERAL INSTITUTE OF TECHNOLOGY, ZURICH for the degree of Doctor of Natural Sciences presented by Carole Andrea Enz Dipl. Natw. ETH Zurich bon1 August 3, 1972 Citizen of Sch(1nholzerswilen (TG), Switzerland accepted on the recommendation of Prof. Dr. J. V. Ward, examiner Prof. Dr. H. Lehtonen, co-examiner Dr. R. Muller, co-examiner Kastanienbaum, 2000 Meinen Eltern und Max Copyright ~;i 2000 by Carole A. Enz, EA WAG Kastanienbaurn All rights reserved. No part of this book rnay be reproduced, stored in a retrieval systen1 or transmitted, in any fonn or by any ineans, electronic, rnechanical, pho- tocopying, recording or otherwise, without the prior written permission of the copyright holder. First Edition 2000 PUBLICATIONS CHAPTER 3 OF THE THESIS HAS BEEN ACCEPTED FOR PUBLICATION: ENZ, C. A., SCHAFFER E. & MOLLER R. Growth and survival of Lake Hallwil whitefish (Co reg onus suidteri) larvae reared on dry and live food. - Archiv fUr Hyclrobiologie. CHAPTERS 4, 5, 6 Ai~D 7 OF THESIS HA VE BEEN SUBMITTED FOR PUBLICATION: ENZ, C. A., MBWENEMO BIA, M. & MULLER. R. Fish species diversity of Lake Hallwil (Switzerland) in the course of eutrophication, with special reference to whitefish ( Coregonus suidteri). Submitted to Conservation Biology. ENZ, C. A .. SCHAFFER, E. & MULLER, R. Importance of prey movement, food particle and tank circulation for rearing Lake Ha11wil whitefish (Coregonus suidteri) larvae. Submitted to North Alnerican Journal of Aquaculture. -

Northern Whitefish (Coregonus Peled) ERSS
U.S. Fish and Wildlife Service Northern Whitefish (Coregonus peled) Ecological Risk Screening Summary U.S. Fish and Wildlife Service, March 2011 Revised, September 2014 and July 2015 Photo not available. 1 Native Range, and Status in the United States Native Range From Froese and Pauly (2015): “Europe and Asia: lakes and rivers from Mezen to Kolyma River, Russia.” Status in the United States This species has not been reported as introduced in the United States. Means of Introductions in the United States This species has not been reported as introduced in the United States. 2 Biology and Ecology Taxonomic Hierarchy and Taxonomic Standing From ITIS (2015): “Kingdom Animalia Subkingdom Bilateria Infrakingdom Deuterostomia Phylum Chordata Subphylum Vertebrata Infraphylum Gnathostomata Superclass Osteichthyes Class Actinopterygii Subclass Neopterygii Infraclass Teleostei Superorder Protacanthopterygii Order Salmoniformes Family Salmonidae Subfamily Coregoninae Genus Coregonus Linnaeus, 1758 – whitefishes Species Coregonus peled (Gmelin, 1789) – peled” “Taxonomic Status: valid” Size, Weight, and Age Range From Froese and Pauly (2015): “Maturity: Lm ?, range 22 - 36 cm Max length : 50.0 cm TL male/unsexed; [Berg 1962]; max. published weight: 5.0 kg [Berg 1962]; max. reported age: 13 years [Kottelat and Freyhof 2007]” Environment From Froese and Pauly (2015): “Marine; freshwater; brackish; demersal; anadromous [Riede 2004].” Climate/Range From Froese and Pauly (2015): “Polar; 74°N - 64°N” Distribution Outside the United States Native From Froese and Pauly (2015): “Europe and Asia: lakes and rivers from Mezen to Kolyma River, Russia.” Introduced From Freyhof and Kottelat (2008): “Hybrids involving C. peled introduced in many reservoirs and lakes (Onega) throughout Russia, eastern and central Europe.” Means of Introduction Outside the United States From Savini et al.