Hymenoptera, Sphecidae) Em Ninhos-Armadilha
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Report on Two Alien Invasive Species of the Genus Sceliphron Klug, 1801 (Hymenoptera Sphecidae) from Sicily, with a Brief Faunistic Update on the Native Species
Biodiversity Journal , 2017, 8 (2): 753–762 A report on two alien invasive species of the genus Sceliphron Klug, 1801 (Hymenoptera Sphecidae) from Sicily, with a brief faunistic update on the native species Giuseppe Fabrizio Turrisi 1,* & Giovanni Altadonna 2 1Via Cristoforo Colombo 8, 95030, Pedara, Catania, Italy; e-mail: [email protected] 2Contrada Filangeri s.n.c., Vill. Pistunina, 98125, Messina, Italy; e-mail: [email protected] *Corresponding author ABSTRACT Two alien invasive species of the genus Sceliphron Klug, 1801 (Hymenoptera Sphecidae) were recently found in Sicily: S. caementarium (Drury, 1773) is recorded from Sicily (Messina province) for the first time; S. curvatum (F. Smith, 1870), previously recorded from Sicily only through generic data from literature and only one locality in a web forum of amateurs, is confirmed as definitely established in the Island. The Regional distribution of both alien species in Italy is revised based on data taken from literature and reliable reports from web forums. A brief faunistic account on the three native Sceliphron from Sicily is provided: S. destillatorium (Illiger, 1807) and S. spirifex (Linnaeus, 1758) are both new for the Aeolian Islands (respectively reported for Panarea and Vulcano). KEY WORDS Sceliphron caementarium ; first record; Sphecidae; Sicily; Italy; alien; invasive species. Received 12.06.2017; accepted 23.06.2017; printed 30.06.2017 INTRODUCTION body with more or less extended yellow spots. The head has a flattened frons, antenna filiform, without In terms of alien species diversity within inver- placoids in the male, distance between antennal tebrate orders, Hymenoptera ranks as third following socket and fronto-clypeal suture at least 0.5 anten - Coleoptera and Hemiptera, with about 300 species, nal socket diameter, mandible without teeth (with representing 30 families, introduced to Europe some exception in the female of a few species) and (Rasplus et al., 2010). -

Last Instar Larva of Penepodium Dubium (Hymenoptera: Sphecidae)
Rev. Biol. Trop., 49(1): 329-332, 2001 www.ucr.ac.cr www.ots.ac.cr www.ots.duke.edu Last instar larva of Penepodium dubium (Hymenoptera: Sphecidae) Sandor Christiano Buys Laboratorio de Entomologia, Departamento de Zoologia, Universidade Federal do Rio de Janeiro, Caixa Postal 68044, CEP 21944-970, Rio de Janeiro, RJ, Brasil, Fax: 55-21-560-5993. email [email protected]. Received 26-VII-1999. Corrected 28-V-2000. Accepted 26-VI-2000. Abstract: The last instar larva of Penepodium dubium is described from southeastern Brazil. This is the first description of an immature stage of the genus Penepodium. The larva of P. dubium is similar to those of Podi- um, but it can be distinguished from the species of this genus by the presence of depressions on the front and clypeus, and lack of brownish blotched areas on the head. The cocoon of Penepodium seems to be unique among those of Sceliphrini in bearing a nipple-like projection at one end. Key words: Penepodium dubium, Sphecidae, Hymenoptera, digger wasp, larva, taxonomy, Brazil. The knowledge on digger wasp larval plastic pots lined inside with absorbent paper morphology has greatly increased since the until the larvae reached maturity. The comprehensive papers of Evans (references in description below is based on three last instar Evans 1964). Even so, the larvae of a great larvae preserved in alcohol (80%), but the number of genera are still unknown. The tribe morphometric features of the head was mea- Sceliphrini is composed by eight genera. The sured in only one specimen. The head and larvae of only four of these genera have been some parts of the integument have been dis- described, namely Chlorion (Evans 1964), sected and heated in KOH (10%) for about 10 Sceliphron (Evans and Lin 1956, Asís et al. -

Guide to Common Tidal Marsh Invertebrates of the Northeastern
- J Mississippi Alabama Sea Grant Consortium MASGP - 79 - 004 Guide to Common Tidal Marsh Invertebrates of the Northeastern Gulf of Mexico by Richard W. Heard University of South Alabama, Mobile, AL 36688 and Gulf Coast Research Laboratory, Ocean Springs, MS 39564* Illustrations by Linda B. Lutz This work is a result of research sponsored in part by the U.S. Department of Commerce, NOAA, Office of Sea Grant, under Grant Nos. 04-S-MOl-92, NA79AA-D-00049, and NASIAA-D-00050, by the Mississippi-Alabama Sea Gram Consortium, by the University of South Alabama, by the Gulf Coast Research Laboratory, and by the Marine Environmental Sciences Consortium. The U.S. Government is authorized to produce and distribute reprints for govern mental purposes notwithstanding any copyright notation that may appear hereon. • Present address. This Handbook is dedicated to WILL HOLMES friend and gentleman Copyright© 1982 by Mississippi-Alabama Sea Grant Consortium and R. W. Heard All rights reserved. No part of this book may be reproduced in any manner without permission from the author. CONTENTS PREFACE . ....... .... ......... .... Family Mysidae. .. .. .. .. .. 27 Order Tanaidacea (Tanaids) . ..... .. 28 INTRODUCTION ........................ Family Paratanaidae.. .. .. .. 29 SALTMARSH INVERTEBRATES. .. .. .. 3 Family Apseudidae . .. .. .. .. 30 Order Cumacea. .. .. .. .. 30 Phylum Cnidaria (=Coelenterata) .. .. .. .. 3 Family Nannasticidae. .. .. 31 Class Anthozoa. .. .. .. .. .. .. .. 3 Order Isopoda (Isopods) . .. .. .. 32 Family Edwardsiidae . .. .. .. .. 3 Family Anthuridae (Anthurids) . .. 32 Phylum Annelida (Annelids) . .. .. .. .. .. 3 Family Sphaeromidae (Sphaeromids) 32 Class Oligochaeta (Oligochaetes). .. .. .. 3 Family Munnidae . .. .. .. .. 34 Class Hirudinea (Leeches) . .. .. .. 4 Family Asellidae . .. .. .. .. 34 Class Polychaeta (polychaetes).. .. .. .. .. 4 Family Bopyridae . .. .. .. .. 35 Family Nereidae (Nereids). .. .. .. .. 4 Order Amphipoda (Amphipods) . ... 36 Family Pilargiidae (pilargiids). .. .. .. .. 6 Family Hyalidae . -

Hymenoptera: Sphecidae: Sphecinae)
The Great Lakes Entomologist Volume 22 Number 4 - Winter 1989 Number 4 - Winter Article 2 1989 December 1989 Distribution and Biology of the Sphecine Wasps of Michigan (Hymenoptera: Sphecidae: Sphecinae) Mark F. O'Brien University of Michigan Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation O'Brien, Mark F. 1989. "Distribution and Biology of the Sphecine Wasps of Michigan (Hymenoptera: Sphecidae: Sphecinae)," The Great Lakes Entomologist, vol 22 (4) Available at: https://scholar.valpo.edu/tgle/vol22/iss4/2 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. O'Brien: Distribution and Biology of the Sphecine Wasps of Michigan (Hymen 1989 THE GREAT LAKES ENTOMOLOGIST 199 DISTRIBUTION AND BIOLOGY OF THE SPHECINE WASPS OF MICHIGAN (HYMENOPTERA: SPHECIDAE: SPHECINAE) Mark F. O'Brien l ABSTRACT Biological information and distribution maps are provided for the 26 species of thread-waisted wasps that occur in Michigan. Podium luctuosum is a new state record. Sixty percent of the eastern North America sphecine fauna is represented in Michigan. The thread-waisted wasps, or Sphecinae (sensu Bohart and Menke 1976) are conspic uous inhabitants of sandy areas, vacant lots, and human residences (mud daubers). They have long been a favorite group for ethologists and naturalists because of their large size and easily observed nesting habits. -

Noaa 13648 DS1.Pdf
r LOAI<CO Qpy N Guide to Gammon Tidal IVlarsh Invertebrates of the Northeastern Gulf of IVlexico by Richard W. Heard UniversityofSouth Alabama, Mobile, AL 36688 and CiulfCoast Research Laboratory, Ocean Springs, MS39564" Illustrations by rimed:tul""'"' ' "=tel' ""'Oo' OR" Iindu B. I utz URt,i',"::.:l'.'.;,',-'-.,":,':::.';..-'",r;»:.",'> i;."<l'IPUS Is,i<'<i":-' "l;~:», li I lb~'ab2 Thisv,ork isa resultofreseaich sponsored inpart by the U.S. Department ofCommerce, NOAA, Office ofSea Grant, underGrani Nos. 04 8 Mol 92,NA79AA D 00049,and NA81AA D 00050, bythe Mississippi Alabama SeaGrant Consortium, byche University ofSouth Alabama, bythe Gulf Coast Research Laboratory, andby the Marine EnvironmentalSciences Consortium. TheU.S. Government isauthorized toproduce anddistribute reprints forgovern- inentalpurposes notwithstanding anycopyright notation that may appear hereon. *Preseitt address. This Handbook is dedicated to WILL HOLMES friend and gentleman Copyright! 1982by Mississippi hlabama SeaGrant Consortium and R. W. Heard All rightsreserved. No part of thisbook may be reproduced in any manner without permissionfrom the author. Printed by Reinbold Lithographing& PrintingCo., BooneviBe,MS 38829. CONTENTS 27 PREFACE FamilyMysidae OrderTanaidacea Tanaids!,....... 28 INTRODUCTION FamilyParatanaidae........, .. 29 30 SALTMARSH INVERTEBRATES ., FamilyApseudidae,......,... Order Cumacea 30 PhylumCnidaria =Coelenterata!......, . FamilyNannasticidae......,... 31 32 Class Anthozoa OrderIsopoda Isopods! 32 Fainily Edwardsiidae. FamilyAnthuridae -

Hymenoptera: Sphecidae: Sphecinae)
The Great Lakes Entomologist Volume 39 Numbers 1 & 2 - Spring/Summer 2006 Numbers Article 14 1 & 2 - Spring/Summer 2006 April 2006 Isodontia Elegans Now in Michigan (Hymenoptera: Sphecidae: Sphecinae) Mark F. O'Brien University of Michigan Julie A. Craves University of Michigan Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation O'Brien, Mark F. and Craves, Julie A. 2006. "Isodontia Elegans Now in Michigan (Hymenoptera: Sphecidae: Sphecinae)," The Great Lakes Entomologist, vol 39 (1) Available at: https://scholar.valpo.edu/tgle/vol39/iss1/14 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. O'Brien and Craves: Isodontia Elegans Now in Michigan (Hymenoptera: Sphecidae: Spheci 94 THE GREAT LAKES ENTOMOLOGIST Vol. 39, Nos. 1 & 2 ISODONTIA ELEGANS NOW IN MICHIGAN (HYMENOPTERA: SPHECIDAE: SPHECINAE) Mark F. O’Brien1 and Julie A. Craves2 ABSTRACT Isodontia elegans (Smith) (Hymenoptera: Sphecidae), one of the “grass- carrying wasps,” previously known only from the western U.S., is now reported from southeastern Michigan, and appears to have spread across the eastern half of the United States in a relatively short period. ____________________ The thread-waisted wasps (Sphecinae) of Michigan are fairly well-studied (O’Brien 1989), with 27 species previously recorded from the state (O’Brien 1989, Cowan 1991). Most of these species are large and easy to identify. -
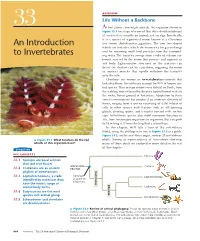
An Introduction to Invertebrates 667 � Figure 33.3 (Continued) Exploring Invertebrate Diversity
OVERVIEW Life Without a Backbone 33 At first glance, you might mistake the organism shown in Figure 33.1 for a type of seaweed. But this colorful inhabitant of coral reefs is actually an animal, not an alga. Specifically, it is a species of segmented worm known as a Christmas An Introduction tree worm (Spirobranchus giganteus). The two tree-shaped whorls are tentacles, which the worm uses for gas exchange and for removing small food particles from the surround- to Invertebrates ing water. The tentacles emerge from a tube of calcium car- bonate secreted by the worm that protects and supports its soft body. Light-sensitive structures on the tentacles can detect the shadow cast by a predator, triggering the worm to contract muscles that rapidly withdraw the tentacles into the tube. Christmas tree worms are invertebrates—animals that lack a backbone. Invertebrates account for 95% of known ani- mal species. They occupy almost every habitat on Earth, from the scalding water released by deep-sea hydrothermal vents to the rocky, frozen ground of Antarctica. Adaptation to these varied environments has produced an immense diversity of forms, ranging from a species consisting of a flat bilayer of cells to other species with features such as silk-spinning glands, pivoting spines, and tentacles covered with suction cups. Invertebrate species also show enormous variation in size, from microscopic organisms to organisms that can grow to 18 m long (1.5 times the length of a school bus). In this chapter, we’ll take a tour of the invertebrate world, using the phylogenetic tree in Figure 33.2 as a guide. -

Abundance and Diversity of Thread-Waisted Wasps (Hymenoptera: Sphecidae: Sphecinae) at Alice L
Transactions of the Illinois State Academy of Science received 7/8/08 (2009), Volume 102, #1&2, pp. 107-115 accepted 1/10/09 Abundance and Diversity of Thread-Waisted Wasps (Hymenoptera: Sphecidae: Sphecinae) at Alice L. Kibbe Life Science Station, Hancock County, Illinois USA Kenneth W. McCravy1*, Jeffrey J. Bara2, Sheri N. Hessler1, Lauren K. Luxmore1, Keesa S. Stinebaker1, and Seán E. Jenkins1 1Department of Biological Sciences, Western Illinois University 1 University Circle, Macomb, IL 61455 2Department of Biology, University of Louisville Louisville, KY 40292 *Corresponding author email: [email protected] ABSTRACT Sphecid wasps (Hymenoptera: Sphecidae) represent a diverse group of predatory wasps that includes potentially important bioindicators and biological control agents. Thread- waisted wasps (Hymenoptera: Sphecidae: Sphecinae) were surveyed in four habitats (recently burned prairie – burned in spring 2005, previously burned prairie – last burned in spring 2004, recently burned oak-hickory forest – burned in spring 2005, and unburned oak-hickory forest – not burned for 5+ years) in west-central Illinois from May through October 2005 using malaise traps. Ten species, representing six genera, were collected. Ammophila nigricans Dahlbom, Eremnophila aureonotata (Cameron), and Sphex nudus Fernald were the most abundant species, representing over 90% of total captures, with A. nigricans alone accounting for almost 75% of all captures. Recently burned prairie pro- duced the greatest overall abundance and species richness, and forest habitats produced the lowest. Relative proportions of sphecine captures in recently burned vs. previously burned prairie decreased late in the season. Greatest numbers of sphecines were captured in July. Seasonal activity patterns were generally similar to those of sphecines further north, but may extend somewhat longer in west-central Illinois for some species. -

FAMILY GROUP NAMES and CLASSIFICATION Superfamily
FAMILY GROUP NAMES AND CLASSIFICATION as of 15 July 2014 compiled by Wojciech J. Pulawski California Academy of Sciences, 55 Music Concourse Drive, Golden Gate Park, San Francisco, CA 94118, USA phone: (415) 379-5313; fax: (415) 379-5715; e-mail: [email protected] The family and tribe level classification used below follows the findings of Brothers (1999), Melo (1999), and Prentice (1998, unpublished Ph.D. thesis, validated by Hanson and Menke, 2006). Their systems differ at the tribal level, and that of Prentice is accepted here as it is based on a much larger data set, but his one new tribe and one new subtribe are not included since they were not published. The classification of Ampulicidae is as proposed by Ohl and Spahn, 2009, the classification of Bembicinae follows Nemkov and Lelej, 2013, and the authorship of Palarini and Xenosphecini is as corrected by Menke and Pulawski (2002). Because Astatinae and Dinetinae each have a single tribe I am not using them in this catalog. The names, authorship, dates, stems, and other pertinent information, are from Menke (1997), who was advised by Don Cameron, a Latin and Greek scholar at the University of Michigan, about the proper formation of names. All genera described subsequent to Bohart and Menke (1976) have been added (both extant and fossil), and recently established synonymies are also indicated. Menke (1997) commented that “Ammophilomorpha, Sphecinomorpha, and Palmodomorpha could be construed as valid [correctly: available] since they are based on generic names with -morpha endings”. Article 11.7.1.3 of the Code (Fourth Edition) leaves no doubt that these names are available, with their original authorship and date, but with a correct suffix (as specified in Article 29.2). -

Hymenoptera: Vespomorpha) Biota Colombiana, Vol
Biota Colombiana ISSN: 0124-5376 [email protected] Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt" Colombia Fernández C., Fernando Checklist of Genera and Subgenera of Aculeate Hymenoptera of the Neotropical Region (Hymenoptera: Vespomorpha) Biota Colombiana, vol. 2, núm. 2, noviembre, 2001, pp. 87- 130 Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt" Bogotá, Colombia Available in: http://www.redalyc.org/articulo.oa?id=49120201 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative FernándezBiota Colombiana 2 (2) 87 - 130, 2001 Himenópteros con Aguijón del Neotrópico -87 Checklist of Genera and Subgenera of Aculeate Hymenoptera of the Neotropical Region (Hymenoptera: Vespomorpha) Fernando Fernández C. Instituto Humboldt A.A. 8693, Bogotá D.C. Colombia. [email protected] Key Words: Aculeate Hymenoptera, Synopsis, Genera, Subgenera, Neotropical Region The aculeate hymenoptera comprises common insects reduction in some groups), and (2) the seventh metasomal like ants, bees and both potter and paper wasps tergum of females is hidden and substantially desclerotized. (Hymenoptera: Vespomorpha or Aculeata). These insects have a significant role in terrestrial ecosystems, especially Superfamily Chrysidoidea in the tropics, acting as pollinators, predators, decomposers, etc. Cuckoo wasps and relatives. This group is basal among the Aculeates. Its phylogeny has been studied by Carpenter The aculeates of the Neotropical Region comprise 3 (1986a, 1999) and Brothers and Carpenter (1993). In the superfamilies, 25 families, 807 genera and about 13000 Neotropical Region it is comprised of 7 families with around described species (Boxes 1 and 2). -

Collection in Linz, Austria Incl
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Linzer biologische Beiträge Jahr/Year: 2016 Band/Volume: 0048_2 Autor(en)/Author(s): Dollfuss Hermann Artikel/Article: The Chloriontinae, Sceliphrinae and Sphecinae Wasps of the "Biologiezentrum Linz"- Collection in Linz, Austria including the Genera Chalybion DAHLBOM, Chlorion LATREILLE, Dynatus LEPELETIER de SAINT FARGEAU, Penepodium MENKE, Podium FABRICIUS, Sceliphron KLUG, Stangeella MENKE and Trigonopsis PERTY (Hymenoptera, Apoidea, Sphecidae) and description of the new species Chalybion ohli from Madagascar 1149-1185 download www.zobodat.at Linzer biol. Beitr. 48/2 1149-1185 19.12.2016 The Chloriontinae, Sceliphrinae and Sphecinae Wasps of the "Biologiezentrum Linz" – Collection in Linz, Austria including the Genera Chalybion DAHLBOM, Chlorion LATREILLE, Dynatus LEPELETIER de SAINT FARGEAU, Penepodium MENKE, Podium FABRICIUS, Sceliphron KLUG, Stangeella MENKE and Trigonopsis PERTY (Hymenoptera, Apoidea, Sphecidae) and description of the new species Chalybion ohli from Madagascar Hermann DOLLFUSS A b s t r a c t : A total of 3.672 specimens of the genera Chalybion (648&&, 908((), Chlorion (40&&, 63((), Dynatus (1(), Penepodium (9&&, 16((), Podium (7&&, 5((), Sceliphron (1.115&&, 822((), Stangeella (12&&, 22((), and Trigonopsis (2&&, 2(() were examined. All information on localities, dates of capture and the name of the collectors are listed. The new species Chalybion ohli from Madagascar and the hitherto unknown males of Chalybion dolichothorax, Chalybion sommereni and the female of Chalybion mochi are described. K e y w o r d s : Apoidea, Sphecidae, Sceliphrinae, Chloriontinae, Chalybion, Chlorion, Dynatus, Penepodium, Podium, Sceliphron, Stangeella, Trigonopsis. Introduction To complete my previous publications (DOLLFUSS 2001, 2004a, 2004b, 2006, 2008a, 2008b, 2010b, 2013b, 2015b) I have identified the Sceliphrini of the collection of "Biologiezentrum Linz" in Austria. -

Nesting Habits, Alternative Nesting Tactics and Female Territoriality of the Cockroach-Hunting Solitary Wasp Penepodium Luteipenne (Hymenoptera, Sphecidae)
JHR 24: 27–41 (2012) Nesting habits, alternative nesting tactics and female territoriality... 27 doi: 10.3897/JHR.24.1828 RESEARCH ARTICLE www.pensoft.net/journals/jhr Nesting habits, alternative nesting tactics and female territoriality of the cockroach-hunting solitary wasp Penepodium luteipenne (Hymenoptera, Sphecidae) Sandor Christiano Buys1 1 Laboratório de Biodiversidade Entomológica, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Avenida Brasil 4.365, Pavilhão Mourisco, sala 201, Manguinhos, 21.045-900, Rio de Janeiro, RJ, Brazil. Corresponding author: Sandor Buys ([email protected]; [email protected]) Academic editor: J. Neff | Received 20 July 2011 | Accepted 27 September 2011 | Published 10 January 2012 Citation: Buys S (2012) Nesting habits, alternative nesting tactics and female territoriality of the cockroach-hunting solitary wasp Penepodium luteipenne (Hymenoptera: Sphecidae). Journal of Hymenoptera Research 24: 27–41. doi: 10.3897/JHR.24.1828 Abstract The nesting behaviour of the cockroach-hunting solitary wasp Penepodium luteipenne (Fabricius, 1804) is described, including five different nesting tactics: digging of new nests, use of pre-existing burrows in the soil, brood parasitism of open nests, brood parasitism of closed nests, and nest usurpation. Observations of a peculiar female territorial behaviour are also provided. Knowledge of the behaviour of other species of the tribe Podiini is summarized and compared with the behaviour of P. luteipenne. This study is based on nesting females observed in the Biological Reserve of Poço das Antas, an area in southeastern Brazil covered by Atlantic Forest. Keywords Behaviour, biology, cleptoparasitism, Podiini Introduction The genus Penepodium Menke, together with the genera Podium Fabricius, Trigonopsis Perty and Dynatus Lepeletier form a monophyletic group of Neotropical cockroach- hunting sphecid wasps (Bohart and Menke 1976, Ohl 1996a, b), currently classified as the tribe Podiini (Pulawski 2011a).