Improved Pastures Support Early Indicators of Soil Restoration in Low-Input Agroecosystems of Nicaragua
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Leveraging Municipal Government Support for Agricultural Value Chains in Nicaragua
LEVERAGING MUNICIPAL GOVERNMENT SUPPORT FOR AGRICULTURAL VALUE CHAINS IN NICARAGUA Leveraging Municipal Government Support for Agricultural Value Chains in Nicaragua Jefferson Shriver and Jorge Manuel Brenes Abdalah HARNESSING THE POWER OF SILC TO DRIVE AGROENTERPRISE DEVELOPMENT IN GHANA Sections of this publication may be copied or adapted to meet local needs without permission from Catholic Relief Services, provided that the parts copied are distributed for free or at cost—not for profit. For any commercial reproduction, please obtain permission from: Catholic Relief Services 228 W. Lexington St. Baltimore, MD 21201 (Tel) 410-625-2220 Attn: PQSD Microfinance Unit (Fax) 410-234-3178 Attn: PQSD Microfinance Unit Email: [email protected] Main CRS website: www.crs.org CRS technical publications: www.crsprogramquality.org Printed in the United States of America Copyright © 2012 Catholic Relief Services ISBN-13: 978-1-61492-111-0 ISBN-10: 1-614-9211-3 HARNESSING THE POWER OF SILC TO DRIVE AGROENTERPRISE DEVELOPMENT IN GHANA Leveraging Municipal Government Support for Agricultural Value Chains in Nicaragua Attracting Private Sector Investment to Rural and Agricultural Markets Jefferson Shriver and Jorge Manuel Brenes Abdalah June 2012 CRACKING THE NUT—ATTRACTING PRIVATE SECTOR INVESTMENT TO RURAL AND AGRICULTURAL MARKETS ACORDAR Project Summary The Alliance to Create Rural Business Opportunities through Agroenterprise Relationships (ACORDAR) is a USAID-sponsored, public-private sector, five- year initiative (2007–12) directly benefitting 7,000 smallholders, organized in 107 cooperatives in 50 municipalities of Nicaragua. Total project investments amount to US$53 million. Project objectives are to assist small producers to increase their income, ensure ongoing employment, and strengthen their market-access capacities, through agricultural value chain development, in alliance with municipal governments, and private sector partnership in Nicaragua. -

Nicaragua: 14 Killed in Anti-Tank Mine Explosion in Jinotega Department Deborah Tyroler
University of New Mexico UNM Digital Repository NotiCen Latin America Digital Beat (LADB) 8-23-1991 Nicaragua: 14 Killed In Anti-tank Mine Explosion In Jinotega Department Deborah Tyroler Follow this and additional works at: https://digitalrepository.unm.edu/noticen Recommended Citation Tyroler, Deborah. "Nicaragua: 14 Killed In Anti-tank Mine Explosion In Jinotega Department." (1991). https://digitalrepository.unm.edu/noticen/5954 This Article is brought to you for free and open access by the Latin America Digital Beat (LADB) at UNM Digital Repository. It has been accepted for inclusion in NotiCen by an authorized administrator of UNM Digital Repository. For more information, please contact [email protected]. LADB Article Id: 065364 ISSN: 1089-1560 Nicaragua: 14 Killed In Anti-tank Mine Explosion In Jinotega Department by Deborah Tyroler Category/Department: General Published: Friday, August 23, 1991 In an Aug. 21 broadcast, Radio Ya reported that 12 recontras under the command of "Indomable" were killed and an indeterminate number wounded when the truck they were traveling in passed over an anti-tank mine, and went over a cliff. The incident occurred near Pita del Carmen, Jinotega department. During an Interior Ministry briefing on Aug. 22, National Police chief Rene Vivas said 14 persons were killed, and seven were recontras. According to Vivas, the mine had intentionally been placed in the roadway, and an investigation was underway. Vivas said seven AK-47 rifles and other materiel were discovered near the site of mine explosion. He added that one of the dead was identified as "El Chino #2," a member of another recontra group headed by "El Tigrillo." Next, in San Ramon, Matagalpa department, a recontra rebel was killed and another wounded in a confrontation with National Police. -
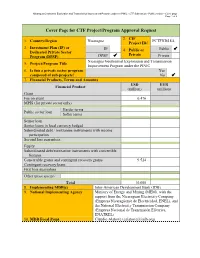
Cover Page for CTF Project/Program Approval Request
Nicaragua Geothermal Exploration and Transmission Improvement Program under the PINIC - CTF Submission - Public version – Cover page Page 1 of 6 Cover Page for CTF Project/Program Approval Request 2. CIF 1. Country/Region Nicaragua PCTFNI618A Project ID# 3. Investment Plan (IP) or IP 4. Public or Public Dedicated Private Sector Private Program (DPSP) DPSP Private Nicaragua Geothermal Exploration and Transmission 5. Project/Program Title Improvement Program under the PINIC 6. Is this a private sector program Yes composed of sub-projects? No 7. Financial Products, Terms and Amounts USD EUR Financial Product (million) (million) Grant Fee on grant 0.476 MPIS (for private sector only) Harder terms Public sector loan Softer terms Senior loan Senior loans in local currency hedged Subordinated debt / mezzanine instruments with income participation Second loss guarantees Equity Subordinated debt/mezzanine instruments with convertible features Convertible grants and contingent recovery grants 9.524 Contingent recovery loans First loss guarantees Other (please specify) Total 10.000 8. Implementing MDB(s) Inter-American Development Bank (IDB) 9. National Implementing Agency Ministry of Energy and Mining (MEM), with the support from the Nicaraguan Electricity Company (Empresa Nicaragüense de Electricidad, ENEL), and the National Electricity Transmission Company (Empresa Nacional de Transmisión Eléctrica, ENATREL) 10. MDB Focal Point Claudio Alatorre ([email protected]) Nicaragua Geothermal Exploration and Transmission Improvement Program under the PINIC - CTF Submission - Public version – Cover page Page 2 of 6 11. Brief Description of Project/Program (including objectives and expected outcomes) In 2015 electricity demand reached 665.4 MW, and it is projected to reach 896 to 1,038 MW by 2026. -

Nicaragua: Chronology of Political Violence & Related Events, February 4 - 10 Deborah Tyroler
University of New Mexico UNM Digital Repository NotiCen Latin America Digital Beat (LADB) 2-14-1992 Nicaragua: Chronology Of Political Violence & Related Events, February 4 - 10 Deborah Tyroler Follow this and additional works at: https://digitalrepository.unm.edu/noticen Recommended Citation Tyroler, Deborah. "Nicaragua: Chronology Of Political Violence & Related Events, February 4 - 10." (1992). https://digitalrepository.unm.edu/noticen/7071 This Article is brought to you for free and open access by the Latin America Digital Beat (LADB) at UNM Digital Repository. It has been accepted for inclusion in NotiCen by an authorized administrator of UNM Digital Repository. For more information, please contact [email protected]. LADB Article Id: 062672 ISSN: 1089-1560 Nicaragua: Chronology Of Political Violence & Related Events, February 4 - 10 by Deborah Tyroler Category/Department: General Published: Friday, February 14, 1992 Feb. 4: Military spokespersons reported one recontra and one recompa killed and four other unidentified combatants wounded during a clash between recontras and recompas near Puerto Viejo, Matagalpa department. Feb. 5: Unidentified assailants burned a bus near Matiguas, Matagalpa. No report on casualties was available. Next, another unidentified group set up a roadblock near Matiguas, forcing five public transport vehicles to stop. One of the vehicles rolled over when the driver attempted to break through the roadblock. Feb. 7: An Interior Ministry spokesperson reported that 346 recontras and 70 recompas surrendered their weapons during a disarmament ceremony held at Tomatoya, Jinotega department. Recontra chiefs "Halcon" and "Culebra" were among the 346. Recontras led by commanders "Peligro," "Xavier," "Punalito," "Saltamontes," and "Terror" agreed to disarm Feb. -

Observing the 2006 Nicaragua Elections
SPECIAL REPORT SERIES Observing the 2006 Nicaragua Elections Waging Peace. Fighting Disease. Building Hope. THE CARTER CENTER STRIVES TO RELIEVE SUFFERING BY ADVANCING PEACE AND HEALTH WORLDWIDE; IT SEEKS TO PREVENT AND RESOLVE CONFLICTS, ENHANCE FREEDOM AND DEMOCRACY, AND PROTECT AND PROMOTE HUMAN RIGHTS WORLDWIDE. OBSERVING THE 2006 NICARAGUA ELECTIONS ONE COPENHILL 453 FREEDOM PARKWAY ATLANTA, GA 30307 (404) 420-5188 FAX (404) 420-5196 WWW.CARTERCENTER.ORG MAY 2007 THE CARTER CENTER OBSERVING THE 2006 NICARAGUA ELECTIONS CONTENTS Foreword . .3 Acknowledgments . .5 Mission Delegation and Staff . 8 Executive Summary . .11 The Pre-election Period . .14 Election Day and Its Aftermath . .32 Conclusions and Recommendations . .39 Appendices . .48 A. Invitation to Observe . .49 B. Role of Long-Term Observers . .51 C. Strategic Deployment of Short-Term Observers . .53 D. Representative Deployment of Short-Term Observers . .58 E. List of Short-Term Deployment Teams . .60 F. Observation Forms . .61 G. Carter Center Public Statements and Press Releases . .67 The Carter Center at a Glance . .90 THE CARTER CENTER OBSERVING THE 2006 NICARAGUA ELECTIONS FOREWORD he Carter Center has monitored national elec- tions in Nicaragua four times, beginning in 1990. Throughout the past 16 years, we have ONTEALEGRE T M sought to ensure that Nicaraguans and the internation- al community were provided with accurate information ARGARITA about the quality of the elections. Carter Center M observers and staff tracked the technical preparations and the electoral campaigns, observed the vote process and count, and provided follow-up visits to monitor electoral justice and the inauguration of new officials. We contributed our knowledge, suggesting to election authorities, political parties, and civil society ways in which the election process could be improved. -
Nicaragua: Government Troops Attack 500 Contras in Chontales Deborah Tyroler
University of New Mexico UNM Digital Repository NotiCen Latin America Digital Beat (LADB) 11-8-1989 Nicaragua: Government Troops Attack 500 Contras In Chontales Deborah Tyroler Follow this and additional works at: https://digitalrepository.unm.edu/noticen Recommended Citation Tyroler, Deborah. "Nicaragua: Government Troops Attack 500 Contras In Chontales." (1989). https://digitalrepository.unm.edu/ noticen/3525 This Article is brought to you for free and open access by the Latin America Digital Beat (LADB) at UNM Digital Repository. It has been accepted for inclusion in NotiCen by an authorized administrator of UNM Digital Repository. For more information, please contact [email protected]. LADB Article Id: 071748 ISSN: 1089-1560 Nicaragua: Government Troops Attack 500 Contras In Chontales by Deborah Tyroler Category/Department: General Published: Wednesday, November 8, 1989 In its Nov. 3 issue, the Sandinista party newspaper Barricada reported that government troops supported by helicopter gunships attacked about 500 contras in La Pinuela and La Cusuca areas of Chontales department. Defense Ministry sources who spoke on condition of anonymity said the army was fighting contra units in the mountains that run from the northern border with Honduras to the southeast. On Nov. 2 the Defense Ministry reported that three contras were killed in El Guapote, 120 miles northeast of the capital in Matagalpa department, and that seven soldiers were wounded during combat in Pantasma and Plan de Grama, Jinotega department. On the same day, Radio Sandino reported that 400 contras who had recently infiltrated into the Quilali area and two other northern districts were "assassinating and kidnapping civilians and making propaganda for the rightist National Opposition Union coalition." (Basic data from AP, 11/03/89) -- End -- ©2011 The University of New Mexico, Latin American & Iberian Institute. -

2020 11 22 USAID-BHA Latin America Storms Fact Sheet #2
Fact Sheet #2 Fiscal Year (FY) 2021 Latin America – Storms NOVEMBER 22, 2020 SITUATION AT A GLANCE 171 5.2 329,700 $69.2 MILLION MILLION Reported Deaths in Latin Estimated People Affected Estimated People in Funding Requested for America due to Eta by Eta and Iota in Central Emergency Shelters in Response to Eta in America Guatemala, Honduras, and Honduras Nicaragua UN – Nov. 19, 2020 UN – Nov. 20, 2020 UN – Nov. 20, 2020 UN – Nov. 19, 2020 Government authorities deliver assistance to storm-affected populations in Colombia’s Providencia and San Andrés islands. Landslides in Guatemala, Honduras, and Nicaragua result in several deaths, with some individuals still missing as of November 20. A The UN issues a $69.2 million appeal for the Honduras humanitarian response to Hurricane Eta’s impacts, while PAHO appeals for $9.9 million to conduct response activities in three countries. TOTAL U.S. GOVERNMENT HUMANITARIAN FUNDING USAID/BHA1 $1,004,808 For the Latin America Storms Response in FY 2021 DoD2 $2,061,681 For complete funding breakdown with partners, see detailed chart on page 6 Total $3,066,426 1 USAID’s Bureau for Humanitarian Assistance (USAID/BHA). Funding figures reflect committed and obligated funding as of November 22, 2020. Total comprises a subset of the more than $17 million in publicly announced USAID/BHA funding to the Latin America and Caribbean storms response 2 U.S. Department of Defense (DoD) 1 TIMELINE KEY DEVELOPMENTS Nov. 3, 2020 Relief Actors Respond to Needs in Colombia’s Eta makes landfall over Providencia and San Andrés Islands Nicaragua’s northeastern coast as a Category 4 Hurricanes Eta and Iota caused widespread damage and destruction in hurricane Colombia’s Providencia and San Andrés islands, including to houses and Nov. -

Latin America – Storms NOVEMBER 22, 2020
Fact Sheet #2 Fiscal Year (FY) 2021 Latin America – Storms NOVEMBER 22, 2020 SITUATION AT A GLANCE 171 5.2 329,700 $69.2 MILLION MILLION Reported Deaths in Latin Estimated People Affected Estimated People in Funding Requested for America due to Eta by Eta and Iota in Central Emergency Shelters in Response to Eta in America Guatemala, Honduras, and Honduras Nicaragua UN – Nov. 19, 2020 UN – Nov. 20, 2020 UN – Nov. 20, 2020 UN – Nov. 19, 2020 Government authorities deliver assistance to storm-affected populations in Colombia’s Providencia and San Andrés islands. Landslides in Guatemala, Honduras, and Nicaragua result in several deaths, with some individuals still missing as of November 20. A The UN issues a $69.2 million appeal for the Honduras humanitarian response to Hurricane Eta’s impacts, while PAHO appeals for $9.9 million to conduct response activities in three countries. TOTAL U.S. GOVERNMENT HUMANITARIAN FUNDING USAID/BHA1 $1,004,808 For the Latin America Storms Response in FY 2021 DoD2 $2,061,681 For complete funding breakdown with partners, see detailed chart on page 6 Total $3,066,426 1 USAID’s Bureau for Humanitarian Assistance (USAID/BHA). Funding figures reflect committed and obligated funding as of November 22, 2020. Total comprises a subset of the more than $17 million in publicly announced USAID/BHA funding to the Latin America and Caribbean storms response 2 U.S. Department of Defense (DoD) 1 TIMELINE KEY DEVELOPMENTS Nov. 3, 2020 Relief Actors Respond to Needs in Colombia’s Eta makes landfall over Providencia and San Andrés Islands Nicaragua’s northeastern coast as a Category 4 Hurricanes Eta and Iota caused widespread damage and destruction in hurricane Colombia’s Providencia and San Andrés islands, including to houses and Nov. -

406947 to 406985—Continued 406947
JANUARY 1 TO DECEMBER 31, 1976 67 406947 to 406985—Continued 406947. ALLIUM CEPA L. Liliaceae. Onion. W-C 1549. One km from Sebaco, Matagalpa Department. White, bulbing onion. Reproduced by seed in Nicaragua. Local name Paisana. Cultivated. Seed. 406948. CAPSICUM ANNUUM L. Solanaceae. Pimento. W-C 1554. Palace Hotel, Managua, Managua Department. Perennial to 2 m tall. Fruit small, red, pungent. Cultivated. Seed. 406949. DIPLADENIA sp. Apocynaceae. W-C 1566. Roadside at El Recreo, Zelaya Department. Slender vine to 2 m tall, leaves opposite, flower salverform, yellow with red eye. Wild. Cuttings. 406950. GOSSYPIUM sp. Malvaceae. W-C 1562. Town of Teustepe, Boaco Department. Shrub to 2 m. Bolls small with 3 or 4 carpels, lint white. Growing both as dooryard and wild cotton. Wild. Seed. 406951. IPOMOEA sp. Convolvulaceae. W-C 1578. Roadside between Esteli and Somoto, Madriz Department. Vigorous twining vine to 3 m long, leafless. Flowers 10 cm in diameter, white. Wild. Cuttings. 406952 to 406968. LYCOPERSICON ESCULENTUM Mill. Solanaceae. Tomato. 406952. W-C 1537. Public market, Managua. Fruit somewhat lobed, red, to 10 cm in diameter. Local name Chilcarojo. Cultivated. Seed. 406953. W-C 1538. Public market, Managua. Pink fruit, catfaced, many lobes, flat, 6 cm by 12 cm. Local name Chilca. Cultivated. Seed. 406954. W-C 1539. Public market, Managua. Large, pear shaped, pink, 5 cm wide by 6 cm long. Local name Roma Chilca. Cultivated. Seed. 406955. W-C 1540. Public market, Managua. Large, pear-to-oblong shaped, lobed at apex, pink, 5 cm by 5 cm. Local name Boca de Mono. Cultivated. -

The Mineral Industry of Nicaragua in 1998
THE MINERAL INDUSTRY OF NICARAGUA By David B. Doan The mineral industry of Nicaragua comprised the production undertaken promptly. Exploration continued in several parts of of moderate but increasing quantities of gold and silver, as well the country. as cement, gypsum, and sand and gravel. A small amount of For more extensive coverage of the mineral industry of refined products were produced for local consumption from Nicaragua, see the 1997 Minerals Yearbook, volume III, The imported oil. Near the end of the year, Hurricane Mitch did Mineral Industries of Latin America and Canada. minor damage to the mining industry, but remediation was THE MINERAL INDUSTRY OF NICARAGUA—1998 20.1 TABLE 1 NICARAGUA: PRODUCTION OF MINERAL COMMODITIES 1/ 2/ (Metric tons unless otherwise specified) Commodity 3/ 1994 1995 e/ 1996 e/ 1997 r/ e/ 1998 e/ Bentonite 704 800 850 926 4/ 875 Cement 308,800 324,000 360,000 r/ 4/ 377,000 4/ 377,000 4/ Gold, mine output, Au content kilograms 1,241 1,316 4/ 1,500 2,562 4/ 3,834 4/ Gypsum and anhydrite, crude 27,900 12,500 12,500 15,820 4/ 22,660 4/ Lime 2,280 3,600 3,600 7,285 4/ 58,527 4/ Petroleum refinery products thousand 42-gallon barrels 4,700 4,800 4,455 r/ 4/ 5,559 4/ 5,650 Salt, marine e/ 15,00 15,000 15,000 13,619 4/ 15,132 4/ Sand and gravel thousand tons 1,200 e/ 1,300 1,350 434 4/ 696 4/ Silver, mine output, Ag content kilograms 2,235 2,400 2,500 1,077 4/ 1,945 4/ e/ Estimated. -

Chagas Disease in Nicaragua
Chagas disease in Nicaragua Implementation of Chagas disease diagnostics for screening and management July 2010 This document is part of a larger case study on Nicaragua’s health system and diagnostic needs. The health system case study employed two main methodological approaches. The first was a review of official Ministry of Health (MINSA) data and epidemiological reports, as well as publications in peer-reviewed journals. The second was a concurrent series of field visits in 2008 to provide firsthand evidence of diagnostic needs and difficulties. A more detailed account of these methodological approaches can be found in the methodology section of the health system case study. Acknowledgment Primary funding for this study was provided by the National Institutes of Health, National Institute of Biomedical Imaging and Bioengineering (grant U54 EB007949). Authors Magda Sequeira, Henry Espinoza, Juan Jose Amador, Gonzalo Domingo, Margarita Quintanilla, and Tala de los Santos, all with PATH (for correspondence: [email protected]). Suggested citation Sequeira M, Espinoza H, Amador JJ, Domingo G, Quintanilla M, and de los Santos T. Chagas Disease in Nicaragua. Seattle, Washington: PATH; 2010. About PATH PATH is an international nonprofit organization that creates sustainable, culturally relevant solutions, enabling communities worldwide to break longstanding cycles of poor health. By collaborating with diverse public- and private-sector partners, PATH helps provide appropriate health technologies and vital strategies that change the way people think and act. PATH’s work improves global health and well-being. Copyright © 2011, Program for Appropriate Technology in Health (PATH). All rights reserved. The material in the document may be freely used for educational or noncommercial purposes, provided that the material is accompanied by an acknowledgment line. -
1 Republic of Nicaragua FY 2019 External Ex-Post Evaluation
Republic of Nicaragua FY 2019 External Ex-Post Evaluation of Japanese Grant Aid Project “The Project for Construction of Paso Real Bridge” External Evaluator: Hajime Sonoda, Global Group 21 Japan, Inc. 0. Summary “The Project for Construction of Paso Real Bridge” (hereinafter referred to as “the Project”) was implemented with the aim of achieving safe and smooth river-crossing traffic over Matagalpa River on National Route 21 by replacing the existing Paso Real Bridge (temporary bridge) located in Matagalpa Department of Nicaragua with a permanent one, thereby contributing to the facilitation of the smooth flow of goods. At the time of both planning and ex-post evaluation, the Project is highly consistent with the development policy/plan and development needs of Nicaragua. As the Project is also consistent with Japan’s assistance policy for Nicaragua at the time of appraisal, the relevance of the Project is high. The outputs of the Project are as planned with both the project cost and project period being within the planned ranges. Therefore, the efficiency of the Project is high. Since the implementation of the Project, there has been no traffic suspensions due to an increased river water level or other reasons at Paso Real Bridge, and the river-crossing time has been shortened. Moreover, the danger posed by over-loaded vehicles above the weight limit crossing the bridge and the number of traffic accidents on the bridge have been reduced. Apart from the fact that the speed limit is not totally adhered to, the purpose of the Project to realize safe and smooth river-crossing traffic has been achieved.