Reconciling the Molecular Clock and Biogeography: an Al- Ternative View of the Divergence Process Between Allognathus Pilsbry, 1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

San Gabriel Chestnut ESA Petition
BEFORE THE SECRETARY OF THE INTERIOR PETITION TO THE U.S. FISH AND WILDLIFE SERVICE TO PROTECT THE SAN GABRIEL CHESTNUT SNAIL UNDER THE ENDANGERED SPECIES ACT © James Bailey CENTER FOR BIOLOGICAL DIVERSITY Notice of Petition Ryan Zinke, Secretary U.S. Department of the Interior 1849 C Street NW Washington, D.C. 20240 [email protected] Greg Sheehan, Acting Director U.S. Fish and Wildlife Service 1849 C Street NW Washington, D.C. 20240 [email protected] Paul Souza, Director Region 8 U.S. Fish and Wildlife Service Pacific Southwest Region 2800 Cottage Way Sacramento, CA 95825 [email protected] Petitioner The Center for Biological Diversity is a national, nonprofit conservation organization with more than 1.3 million members and supporters dedicated to the protection of endangered species and wild places. http://www.biologicaldiversity.org Failure to grant the requested petition will adversely affect the aesthetic, recreational, commercial, research, and scientific interests of the petitioning organization’s members and the people of the United States. Morally, aesthetically, recreationally, and commercially, the public shows increasing concern for wild ecosystems and for biodiversity in general. 1 November 13, 2017 Dear Mr. Zinke: Pursuant to Section 4(b) of the Endangered Species Act (“ESA”), 16 U.S.C. §1533(b), Section 553(3) of the Administrative Procedures Act, 5 U.S.C. § 553(e), and 50 C.F.R. §424.14(a), the Center for Biological Diversity and Tierra Curry hereby formally petition the Secretary of the Interior, through the United States Fish and Wildlife Service (“FWS”, “the Service”) to list the San Gabriel chestnut snail (Glyptostoma gabrielense) as a threatened or endangered species under the Endangered Species Act and to designate critical habitat concurrently with listing. -

Biogeography of the Land Snail Genus Allognathus (Helicidae): Middle Miocene Colonization of the Balearic Islands
Journal of Biogeography (J. Biogeogr.) (2015) 42, 1845–1857 ORIGINAL Biogeography of the land snail genus ARTICLE Allognathus (Helicidae): middle Miocene colonization of the Balearic Islands Luis J. Chueca1,2*, Marıa Jose Madeira1,2 and Benjamın J. Gomez-Moliner 1,2 1Department of Zoology and Animal Cell ABSTRACT Biology, Faculty of Pharmacy, University of Aim We infer the evolutionary history of the land snail genus Allognathus the Basque Country, 01006 Vitoria-Gasteiz, 2 from a molecular phylogeny. An approximate temporal framework for its colo- Alava, Spain, Biodiversity Research Group CIEA Lucio Lascaray, 01006 Vitoria-Gasteiz, nization of the Balearic Islands and diversification within the archipelago is Alava, Spain provided according to palaeogeographical events in the western Mediterranean Basin. Location The Balearic Islands, Western Mediterranean. Methods A 2461-bp DNA sequence dataset was generated from one nuclear and two mitochondrial gene fragments in 87 specimens, covering all nominal taxa of the genus Allognathus. Through maximum-likelihood and Bayesian phylogenetic methods along with a Bayesian molecular clock, we examined the evolutionary history of the group. Ancestral distribution ranges were estimated for divergence events across the tree using a Bayesian approach. We also used genetic species-delimitation models to determine the taxonomy of Allognathus. Results We provided the first molecular phylogeny of Allognathus, a genus endemic to the Balearic Islands. The origin of the genus in the Balearic Islands was dated to the middle Miocene based on palaeogeographical events in the Western Mediterranean. During the late Miocene and Pliocene, several diversi- fication events occurred within the archipelago. The ancestral range of Allogna- thus was reconstructed as the north-eastern Tramuntana Mountains of Mallorca. -

Compositional Variability of Pleistocene Land Snail Assemblages Preserved in a Cinder Cone
Compositional variability of Pleistocene land snail assemblages preserved in a cinder cone volcano from Tenerife, Canary Islands A thesis submitted to the graduate school of the University of Cincinnati in partial fulfillment of the requirements for the degree of Master of Science In the Department of Geology of the College of Arts and Sciences by Elizabeth M. Bullard B.S., Muskingum University, 2012 July 2016 Advisors: Dr. Yurena Yanes Dr. Arnold I. Miller Committee Member: Dr. Joshua Miller i Abstract Fossil assemblage faunal compositions may vary through space and time in response to climatic and/or taphonomic factors, but these relationships can be difficult to diagnose and disentangle. Here, we investigate how to disentangle climatic and taphonomic signals of a land- snail-rich volcanic scoria sequence to asses if it was influenced by taphonomic bias, climate change, or both, using a multifaceted approach, combining taphonomic, ecological, body-size, and stable-isotope data. Fossil assemblages were sampled from two beds (Units A and B) in a Pleistocene cinder cone volcano of southern Tenerife (Canary Islands), dated to the glacial interval MIS 8 (~299-302 ka). The two units differed in taphonomy, species composition, and abundance distributions. The upper unit, B (6 species), showed higher snail diversity and shell concentration and lower taphonomic alteration than the lower unit, A (3 species). Furthermore, larger bodied species (length>10mm) dominated Unit A and were better preserved than smaller species (length<10mm), whereas smaller individuals were more abundant (and better preserved) at Unit B. These differences were likely impacted by physical differences in the matrices surrounding the fossils. -

Diplom-Biologe KLAUS GROH Malakozoologe Und Naturschützer – 65 Jahre
53 Mitt. dtsch. malakozool. Ges. 94 53 – 70 Frankfurt a. M., November 2015 Diplom-Biologe KLAUS GROH Malakozoologe und Naturschützer – 65 Jahre CARSTEN RENKER & JÜRGEN H. JUNGBLUTH th Abstract: The 65 birthday of KLAUS GROH is a good occasion to give a retrospect of his life and hitherto existing achievement. Beside his vita we summarize his malacological work, give an overview about the projects for the protection of species, have a look on his tremendous impetus for the worldwide distribution of malacological knowledge by the establishment of the CHRISTA HEMMEN-Verlag, later ConchBooks, as publishing house, book trader and antiquarian. Last but not least we give a summary of his scientific achievements culminating in 206 publications and containing descriptions of up to now 42 specific taxa. Keywords: KLAUS GROH, biography, bibliography, malacology, freshwater mussels, Hesse, Rhineland- Palatinate, Luxembourg Zusammenfassung: Der 65. Geburtstag von KLAUS GROH wird zum Anlass genommen einen Rückblick auf sein bisheriges Leben und Wirken zu geben. Neben der Vita werden vor allem seine malakologische Arbeit und sein ehrenamtliches Engagement in zahlreichen malakologischen Verbänden und Naturschutzvereinen betrachtet. KLAUS GROH nahm außerdem einen enormen Einfluss auf die weltweite Verbreitung malako- logischen Wissens durch die Gründung des CHRISTA HEMMEN-Verlags, später ConchBooks, als Verlagshaus, Buchhandlung und Antiquariat. Schließlich gilt es seine wissenschaftlichen Verdienste zu würdigen, die in 206 Publikationen und Neubeschreibungen 42 spezifischer Taxa kulminieren. Vita Schulzeit Am 22. Mai 1949 wurde KLAUS GROH in Darmstadt als Sohn des Bauschlossers HELMUT GROH und seiner Ehefrau ANNELIESE, geb. FEDERLEIN geboren. Er besuchte die Volksschulen in Langen/Hessen und Kirchheim unter Teck/Baden-Württemberg (1955-1959), es folgte der Besuch der Realschule in Langen/Hessen (1959-1965), dort schloss er auch seine Schulzeit mit der „Mittleren Reife“ ab. -

Description of a Representative of the Genus Hemicycla Canary Islands
BASTERIA, 70: 53-56, 2006 of discovered extinct of the Description a newly representative genus Hemicycla Swainson, 1840 (Gastropoda, Pulmonata, Helicidae) from La Gomera, Canary Islands T. Beck Institut fur Palaontologie der Universitat Erlangen, Loewenichstrasse 28, D 91054 Erlangen, Germany; [email protected] & W. Rähle Engelfriedshalde 102, D-72076 Tubingen, Germany; [email protected] A fossil of the from La Gomera is described. It found in new species genus Hemicycla was Quaternary deposits near Hermigua in the northeast of the island. The shells are considerably larger than those of all other Hemicycla species from the Canary Islands. Hemicycla montefortia- shows malleated shell whereas the shell surface smooth. na spec. nov. a strongly sculpture is Key words: Gastropoda, Pulmonata,Helicidae, Hemicycla, taxonomy, new species, Quaternary, La Gomera, Canary Islands, Spain. At the slopes of numerous barrancos on the island of La Gomera Quaternary depo- sits are exposed. They are deeply cut by the erosive activities of the actual brooks. According to Groh et al. (1996) these deposits have to be interpreted as remnants of the bottoms of ancient, presumably Pleistocene valleys. At many localities shells of land snails can be found in such deposits. In some out- that still the island with crops species are living on are co-occuring supposed extinct spe- cies only known from fossil records. In the checklist Bank, Groh & 12 of the published by Ripken (2002) species genus Hemicycla Swainson, 1840, are mentioned to occur on La Gomera. Five taxa [J[H. gomerensis (Mousson, 1872), H. hedybia (Mabille, 1882), H. paivanopsis (Mabille, 1882), H. -

Hemicycla (Adiverticula) Diegoi (Gastropoda: Pulmonata: Helicidae
Zootaxa 2757: 29–46 (2011) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2011 · Magnolia Press ISSN 1175-5334 (online edition) Hemicycla (Adiverticula) diegoi (Gastropoda: Pulmonata: Helicidae), a new species from Tenerife, Canary Islands, with a phylogenetic analysis of conchologically similar species in the genus Hemicycla Swainson, 1840 MARCO T. NEIBER1,5, RICARDO VEGA-LUZ2, RODOLFO VEGA-LUZ3 & STEFAN KOENEMANN4 1University of Veterinary Medicine Hannover, Institute for Animal Ecology and Cell Biology, Bünteweg 17d, D-30559 Hannover, Ger- many. E-mail: [email protected] 2C/ Golf de El Candido 49, E-29018 Malaga, Spain. E-mail: [email protected] 3Ctra. Punta de Teno s/n., Teno Bajo, E-38480 Buenavista del Norte, Spain 4University of Siegen, Department of Biology and Didactics, Adolf-Reichwein-Str. 2, D-57068 Siegen, Germany. E-mail: koene- [email protected] 5 Corresponding author Abstract Hemicycla (Adiverticula) diegoi n. sp. from the westernmost part of the Teno massif above the Lighthouse at Punta de Teno, Tenerife, Canary Islands is described and compared to conchologically and anatomically similar species in the genus Hemicycla Swainson, 1840. The validity of the new species is corroborated by a phylogenetic analysis including several congeneric species, inter- and intraspecific genetic distances and a morphometric comparison by means of a discriminant function analysis. Hemicycla cf. paivanopsis (Mabille, 1882) and Hemicycla quadricincta quadricincta (Morelet, 1864) from La Gomera and Hemicycla berkeleii (R. T. Lowe, 1861) from Gran Canaria are tentatively placed in Hemicycla s. str. on the basis of a phylogenetic analysis. Furthermore, Eobania vermiculata (O. -
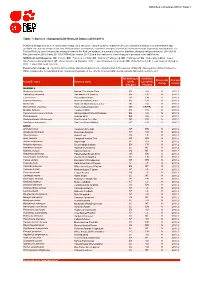
Table 7: Species Changing IUCN Red List Status (2010-2011)
IUCN Red List version 2011.2: Table 7 Table 7: Species changing IUCN Red List Status (2010-2011) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2010 (IUCN Red List version 2010.4) and 2011 (IUCN Red List version 2011.2) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.) IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2010) List (2011) change version Category Category MAMMALS Bradypus torquatus Maned Three-toed Sloth EN VU N 2011.1 Callicebus oenanthe San Martin Titi Monkey EN CR N 2011.1 Equus ferus Przewalski's Horse CR EN G 2011.2 -

Una Nueva Especie De Tambja Burn, 1962 (Mollusca, Nudibranchia)
Bull. Mus. natn. Hist. nal.. Paris. 4' sér., 10, 1988, section A, n° 2 : 301-307. Una nueva especie de Tambja Burn, 1962 (Mollusca, Nudibranchia) por José Carlos GARCÍA GÓMEZ y Jesús ORTEA Resumen. — Una nueva especie de Tambja Burn, 1962 del estrecho de Gibraltar, Tambja ceutae n. sp. se distingue de todas las restantes especies del género por la presencia de pequeñas papilas cónicas bordeando el noto, y por su próstata netamente diferenciada del canal deferente. Su notable coloración, verde azulada con bandas longitudinales amarillas, hace que sea uno de los Nudibranquios mas espectaculares del área europea, donde no puede ser confundido con ningún otro. Resume. — Une nouvelle espèce de Tambja Burn, 1962 du détroit de Gibraltar, Tambja ceutae n. sp., se distingue de toutes les autres espèces du genre par la présence de petites papilles coniques bordant le notum, et par sa prostate nettement différenciée du canal déférent. Sa couleur remarquable, bleu vert avec des bandes longitudinales jaunes, en fait un des Nudibranches les plus spectaculaires du domaine européen, où il ne peut être confondu avec aucun autre. J. C. GARCÍA GÓMEZ, Dep. Zoología, Fac. Biología, Univ. Sevilla, España. ¡. ORTEA, Dep. Zoología, Fac. Biología, Univ. Oviedo, España. INTRODUCCIÓN En un trabajo anterior, ORTEA y GARCÍA (1986), describíamos una nueva especie de Tambja Burn, capturada en las islas de Cabo Verde, que suponía la primera referencia en el Atlántico Nordeste de un género de Gymnodoridos sin procesos frontales cuya rádula se caracteriza por tener una muesca en la zona media del diente raquídeo y un primer diente lateral bífido. -

Gastropoda, Pulmonata, Helicidae): H
Journal of Conchology (2009), Vol.40, No.1 31 REDISCOVERY AND REDESCRIPTION OF AN ALMOST UNKNOWN HEMICYCLA SPECIES (GASTROPODA, PULMONATA, HELICIDAE): H. EURYTHYRA O. BOETTGER 1908 FROM TENERIFE, CANARY ISLANDS Yurena Yanes1, Javier Martín2, Miguel Artiles3, Leopoldo Moro4, María R. Alonso5 & Miguel Ibáñez5 1Savannah River Ecology Laboratory, the University of Georgia, Drawer E, Aiken SC 29802 2El Coromoto, La Laguna, Tenerife, Canary Islands, Spain 3Arinaga, Gran Canaria, Canary Islands, Spain 4Centro de Planificación Ambiental (CEPLAM). Tenerife, Canary Islands, Spain 5Department of Animal Biology, La Laguna University, E–38206 La Laguna, Tenerife, Canary Islands, Spain Abstract Hemicycla eurythyra O. Boettger 1908, endemic to Tenerife, was rediscovered near its type locality. It is rede- scribed and compared with two conchologically similar taxa: H. pouchet (A. Férussac 1821) and H. pouchadan Ibáñez & Alonso 2007. H. eurythyra is grouped in the subgenus Hemicycla s. str. by the presence of a diverticulum in the bursa copulatrix complex. Key words Gastropoda, Hemicycla eurythyra, Tenerife, Canary Islands. INTRODUCT I ON J. Martín. It consists of ten living specimens from Bajamar (leg. M. Artiles, May 2008) and several Boettger (1908) made a brief conchological des- fresh and Quaternary fossil shells from the area cription of a Quaternary fossil taxon from the shown in Fig. 1, collected between 1985 and Tejina Ravine, Tenerife Island, Canary Archipelago 2007. The fossil shells come from a Pleistocene (Fig. 1), with the name Hemicycla puchet var. aeolian deposit older than 130 kyr BP (Castillo eurythyra, as follows: et al., 2006). The photographic methodology used is described in detail by Ibáñez et al. (2006). -

Contribución Al Conocimiento De Los Moluscos Fósiles De Las Islas Canarias
SPIRA 2008 Vol. 2 Núm. 4 Pàg. 199-221 Rebut el 19 de juliol de 2007. Acceptat el 25 de gener de 2008 Contribución al conocimiento de los moluscos fósiles de las Islas Canarias JULIO TALAVÁN SERNA & JULIO TALAVÁN GÓMEZ Valle de la Ballestera 32-16, 46015 València E-mail: [email protected] Resumen.—Contribución al conocimiento de los moluscos fósiles de las Islas Canarias. En este artículo se presenta una breve relación de algunas especies fósiles y subfósiles de moluscos continentales de las Islas Canarias, aportando además comentarios acerca de la distribución, el hábitat y el estatus taxonómico de muchas de ellas. Palabras clave.—Mollusca, Continental, Fósiles, Islas Canarias, España. Resum.—Contribució al coneixement dels mol·luscos fòssils de les Illes Canàries. En aquest article es presenta una breu relació d´algunes espècies fòssils i subfòssils de mol·luscos continentals de les Illes Canàries, aportant a més comentaris sobre la distribució, l’hàbitat i l’estatus taxonòmic de moltes d’elles. Paraules clau.—Mollusca, Continental, Fòssils, Illes Canàries, Espanya. Abstract.—Contribution to the knowledge of fossil mollusks from the Canary Islands. In this paper a brief account of some fossil and subfossil species of continental mollusks from the Canary Islands is reported, further including comments on the distribution, habitat and taxonomic status of many of them. Key words.—Mollusca, Continental, Fossils, Canary Islands, Spain. _______________________________________________________________________________________________ INTRODUCCIÓN A lo largo de diferentes años, desde 1996 a 2006, hemos realizado varios viajes El archipiélago canario se encuentra a estas islas, aprovechando el periodo situado apenas a 100 km al oeste de la estival, durante los cuales hemos recogido costa africana y a unos 400 km del trópico el material que constituye la base de este de Cáncer. -

A Phylogenetic Analysis of Polygyridae (Gastropoda: Pulmonata) Based on Mitochondrial DNA Sequence Data Nicholas A
Southern Illinois University Carbondale OpenSIUC Honors Theses University Honors Program 2012 A Phylogenetic Analysis of Polygyridae (Gastropoda: Pulmonata) Based on Mitochondrial DNA Sequence Data Nicholas A. Defreitas Southern Illinois University Carbondale, [email protected] Follow this and additional works at: http://opensiuc.lib.siu.edu/uhp_theses Recommended Citation Defreitas, Nicholas A., "A Phylogenetic Analysis of Polygyridae (Gastropoda: Pulmonata) Based on Mitochondrial DNA Sequence Data" (2012). Honors Theses. Paper 348. This Dissertation/Thesis is brought to you for free and open access by the University Honors Program at OpenSIUC. It has been accepted for inclusion in Honors Theses by an authorized administrator of OpenSIUC. For more information, please contact [email protected]. A Phylogenetic Analysis of Polygyridae (Gastropoda: Pulmonata) Based on Mitochondrial DNA Sequence Data Nicholas Defreitas University Honors Program Senior Thesis Introduction Despite the increasing use of molecular methods to determine evolutionary relationships among taxa, molecular sequence data have never been used to assess the relationships among the polygyrid snails (Gastropoda:Pulmonata:Polygyridae). This is surprising, considering how large, charismatic and common they are. Polygyrids range across North America, going as far north as parts of Canada and south as Mexico and even deeper into Central America (Pilsbry 1940). There is a particular concentration of these snails in the Appalachian Mountains, where they primarily serve as detritivores and prey for various woodland vertebrates in forest habitats. Yet despite the broad geographic distribution and high abundance of polygyrids in many forest habitats, there is still little known about their phylogeny (evolutionary relationships). Polygyrids are broadly distributed across North America. Mesodontini and Triodopsini are both found in eastern North America (Hubricht 1985). -

Folia Conchyliologica 60 Altab
An enigmatic rock-dwelling land snail from southernmost Iberia (Mollusca, Gastropoda, Helicidae) Cristian Altaba, Félix Jiménez To cite this version: Cristian Altaba, Félix Jiménez. An enigmatic rock-dwelling land snail from southernmost Iberia (Mollusca, Gastropoda, Helicidae). Folia Conchyliologica, Cédric Audibert, 2021, pp.35-44. hal- 03144942 HAL Id: hal-03144942 https://hal.archives-ouvertes.fr/hal-03144942 Submitted on 24 Feb 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Folia Conchyliologica, 60 Février 2021 An enigmatic ro ck - dwelling land snail from southernmost Iberia (Mollusca, Gastropoda, Helicidae) 1,2 3 Cristian R. ALTABA & Félix RÍOS JIMÉNEZ 1 Llevant Natural Park, Department of the Environm ent, Government of the Balearic Islands, Ap. 76, 07570 Artà, Balearic Island s, Spain. 2 Research Group on Human Evolution and Cogniton (EVOCOG), University of the Balearic Islands, 07072 Palma, Balearic Islands, Spain; [email protected] (correspondi ng author). 3 Sociedad Gaditana de Historia Natural, c/Madreselva s/n, 11408 Jerez, Cádiz, Spain. http://zoobank.org/References/7dab39f0 - 43d3 - 4804 - 976c - 71a1de49f801 Abstract - A new species and genus of helicid land snail, Tartessiberus cilbanus , is herein described from the extreme south of continental Spain.