The Relative Rates of Thiol–Thioester Exchange and Hydrolysis for Alkyl and Aryl Thioalkanoates in Water
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Alkyldihydropyrones, New Polyketides Synthesized by a Type III Polyketide Synthase from Streptomyces Reveromyceticus
The Journal of Antibiotics (2014) 67, 819–823 & 2014 Japan Antibiotics Research Association All rights reserved 0021-8820/14 www.nature.com/ja ORIGINAL ARTICLE Alkyldihydropyrones, new polyketides synthesized by a type III polyketide synthase from Streptomyces reveromyceticus Teruki Aizawa1, Seung-Young Kim1, Shunji Takahashi2,3, Masahiko Koshita1, Mioka Tani1, Yushi Futamura3, Hiroyuki Osada2,3 and Nobutaka Funa1 Genome sequencing allows a rapid and efficient identification of novel catalysts that produce novel secondary metabolites. Here we describe the catalytic properties of dihydropyrone synthase A (DpyA), a novel type III polyketide synthase encoded in a linear plasmid of Streptomyces reveromyceticus. Heterologous expression of dpyA led to the accumulation of alkyldihydropyrones A (1), B (2), C (3) and D (4), which are novel dihydropyran compounds that exhibit weak cytotoxicity against the leukemia cell line HL-60. DpyA catalyzes the condensation of b-hydroxyl acid thioester and methylmalonyl-CoA to yield a triketide intermediate that then undergoes lactonization of a secondary alcohol and a thioester to give alkyldihydropyrone. The Journal of Antibiotics (2014) 67, 819–823; doi:10.1038/ja.2014.80; published online 2 July 2014 INTRODUCTION Very recently, Tang et al.11 discovered that presulficidin, which is Type III polyketide synthase (PKS), which is widely distributed synthesized by Cpz6, a type III PKS from the caprazamycin among higher plants, fungi and bacteria, catalyzes the assembly of biosynthetic cluster, relays sulfonate from 30-phosphoadenine-50- primary metabolites such as acyl-CoA compounds to synthesize phosphosulfate to caprazamycin. structurally complex polyketides.1 The reaction catalyzed by type III Genome sequence analyses of Streptomyces species have shown that PKS starts with the formation of a thioester bond between the the number of biosynthetic gene clusters encoded on the chromosome catalytic center cysteine and an acyl group derived from a starter is much higher than the number of secondary metabolites isolated substrate. -

The Reaction of Aminonitriles with Aminothiols: a Way to Thiol-Containing Peptides and Nitrogen Heterocycles in the Primitive Earth Ocean
life Article The Reaction of Aminonitriles with Aminothiols: A Way to Thiol-Containing Peptides and Nitrogen Heterocycles in the Primitive Earth Ocean Ibrahim Shalayel , Seydou Coulibaly, Kieu Dung Ly, Anne Milet and Yannick Vallée * Univ. Grenoble Alpes, CNRS, Département de Chimie Moléculaire, Campus, F-38058 Grenoble, France; [email protected] (I.S.); [email protected] (S.C.); [email protected] (K.D.L.); [email protected] (A.M.) * Correspondence: [email protected] Received: 28 September 2018; Accepted: 18 October 2018; Published: 19 October 2018 Abstract: The Strecker reaction of aldehydes with ammonia and hydrogen cyanide first leads to α-aminonitriles, which are then hydrolyzed to α-amino acids. However, before reacting with water, these aminonitriles can be trapped by aminothiols, such as cysteine or homocysteine, to give 5- or 6-membered ring heterocycles, which in turn are hydrolyzed to dipeptides. We propose that this two-step process enabled the formation of thiol-containing dipeptides in the primitive ocean. These small peptides are able to promote the formation of other peptide bonds and of heterocyclic molecules. Theoretical calculations support our experimental results. They predict that α-aminonitriles should be more reactive than other nitriles, and that imidazoles should be formed from transiently formed amidinonitriles. Overall, this set of reactions delineates a possible early stage of the development of organic chemistry, hence of life, on Earth dominated by nitriles and thiol-rich peptides (TRP). Keywords: origin of life; prebiotic chemistry; thiol-rich peptides; cysteine; aminonitriles; imidazoles 1. Introduction In ribosomes, peptide bonds are formed by the reaction of the amine group of an amino acid with an ester function. -

Chapter 7, Haloalkanes, Properties and Substitution Reactions of Haloalkanes Table of Contents 1
Chapter 7, Haloalkanes, Properties and Substitution Reactions of Haloalkanes Table of Contents 1. Alkyl Halides (Haloalkane) 2. Nucleophilic Substitution Reactions (SNX, X=1 or 2) 3. Nucleophiles (Acid-Base Chemistry, pka) 4. Leaving Groups (Acid-Base Chemistry, pka) 5. Kinetics of a Nucleophilic Substitution Reaction: An SN2 Reaction 6. A Mechanism for the SN2 Reaction 7. The Stereochemistry of SN2 Reactions 8. A Mechanism for the SN1 Reaction 9. Carbocations, SN1, E1 10. Stereochemistry of SN1 Reactions 11. Factors Affecting the Rates of SN1 and SN2 Reactions 12. --Eliminations, E1 and E2 13. E2 and E1 mechanisms and product predictions In this chapter we will consider: What groups can be replaced (i.e., substituted) or eliminated The various mechanisms by which such processes occur The conditions that can promote such reactions Alkyl Halides (Haloalkane) An alkyl halide has a halogen atom bonded to an sp3-hybridized (tetrahedral) carbon atom The carbon–chlorine and carbon– bromine bonds are polarized because the halogen is more electronegative than carbon The carbon-iodine bond do not have a per- manent dipole, the bond is easily polarizable Iodine is a good leaving group due to its polarizability, i.e. its ability to stabilize a charge due to its large atomic size Generally, a carbon-halogen bond is polar with a partial positive () charge on the carbon and partial negative () charge on the halogen C X X = Cl, Br, I Different Types of Organic Halides Alkyl halides (haloalkanes) sp3-hybridized Attached to Attached to Attached -

Identification of a Thioesterase Bottleneck in the Pikromycin Pathway Through Full-Module Processing of Unnatural Pentaketides
Article pubs.acs.org/JACS Identification of a Thioesterase Bottleneck in the Pikromycin Pathway through Full-Module Processing of Unnatural Pentaketides † ‡ † § † ‡ ⊥ ∥ Douglas A. Hansen, , Aaron A. Koch, , and David H. Sherman*, , , , † ‡ § ⊥ Life Sciences Institute, Department of Medicinal Chemistry, Cancer Biology Graduate Program, Department of Chemistry, and ∥ Department of Microbiology & Immunology, University of Michigan, Ann Arbor, Michigan 48109, United States *S Supporting Information ABSTRACT: Polyketide biosynthetic pathways have been engineered to generate natural product analogs for over two decades. However, manipulation of modular type I polyketide synthases (PKSs) to make unnatural metabolites commonly results in attenuated yields or entirely inactive pathways, and the mechanistic basis for compromised production is rarely elucidated since rate-limiting or inactive domain(s) remain unidentified. Accordingly, we synthesized and assayed a series of modified pikromycin (Pik) pentaketides that mimic early pathway engineering to probe the substrate tolerance of the PikAIII-TE module in vitro. Truncated pentaketides were processed with varying efficiencies to corresponding macrolactones, while pentaketides with epimerized chiral centers were poorly processed by PikAIII-TE and failed to generate 12-membered ring products. Isolation and identification of extended but prematurely offloaded shunt products suggested that the Pik thioesterase (TE) domain has limited substrate flexibility and functions as a gatekeeper in the processing of -

Nomenclature of Carboxylic Acids • Select the Longest Carbon Chain Containing the Carboxyl Group
Chapter 5 Carboxylic Acids and Esters Carboxylic Acids • Carboxylic acids are weak organic acids which Chapter 5 contain the carboxyl group (RCO2H): Carboxylic Acids and Esters O C O H O RCOOH RCO2H Chapter Objectives: O condensed ways of • Learn to recognize the carboxylic acid, ester, and related functional groups. RCOH writing the carboxyl • Learn the IUPAC system for naming carboxylic acids and esters. group a carboxylic acid C H • Learn the important physical properties of the carboxylic acids and esters. • Learn the major chemical reaction of carboxylic acids and esters, and learn how to O predict the products of ester synthesis and hydrolysis reactions. the carboxyl group • Learn some of the important properties of condensation polymers, especially the polyesters. Mr. Kevin A. Boudreaux • The tart flavor of sour-tasting foods is often caused Angelo State University CHEM 2353 Fundamentals of Organic Chemistry by the presence of carboxylic acids. Organic and Biochemistry for Today (Seager & Slabaugh) www.angelo.edu/faculty/kboudrea 2 Nomenclature of Carboxylic Acids • Select the longest carbon chain containing the carboxyl group. The -e ending of the parent alkane name is replaced by the suffix -oic acid. • The carboxyl carbon is always numbered “1” but the number is not included in the name. • Name the substituents attached to the chain in the Nomenclature of usual way. • Aromatic carboxylic acids (i.e., with a CO2H Carboxylic Acids directly connected to a benzene ring) are named after the parent compound, benzoic acid. O C OH 3 -

Introduction to Alkenes and Alkynes in an Alkane, All Covalent Bonds
Introduction to Alkenes and Alkynes In an alkane, all covalent bonds between carbon were σ (σ bonds are defined as bonds where the electron density is symmetric about the internuclear axis) In an alkene, however, only three σ bonds are formed from the alkene carbon -the carbon thus adopts an sp2 hybridization Ethene (common name ethylene) has a molecular formula of CH2CH2 Each carbon is sp2 hybridized with a σ bond to two hydrogens and the other carbon Hybridized orbital allows stronger bonds due to more overlap H H C C H H Structure of Ethylene In addition to the σ framework of ethylene, each carbon has an atomic p orbital not used in hybridization The two p orbitals (each with one electron) overlap to form a π bond (p bonds are not symmetric about the internuclear axis) π bonds are not as strong as σ bonds (in ethylene, the σ bond is ~90 Kcal/mol and the π bond is ~66 Kcal/mol) Thus while σ bonds are stable and very few reactions occur with C-C bonds, π bonds are much more reactive and many reactions occur with C=C π bonds Nomenclature of Alkenes August Wilhelm Hofmann’s attempt for systematic hydrocarbon nomenclature (1866) Attempted to use a systematic name by naming all possible structures with 4 carbons Quartane a alkane C4H10 Quartyl C4H9 Quartene e alkene C4H8 Quartenyl C4H7 Quartine i alkine → alkyne C4H6 Quartinyl C4H5 Quartone o C4H4 Quartonyl C4H3 Quartune u C4H2 Quartunyl C4H1 Wanted to use Quart from the Latin for 4 – this method was not embraced and BUT has remained Used English order of vowels, however, to name the groups -

Linear Alkylbenzene Sulphonate (CAS No
Environmental Risk Assessment LAS Linear Alkylbenzene Sulphonate (CAS No. 68411-30-3) Revised ENVIRONMENTAL Aspect of the HERA Report = February 2013 = 1 1. Contents 2. Executive summary 3. Substance characterisation 3.1 CAS No. and grouping information 3.2 Chemical structure and composition 3.3 Manufacturing route and production/volume statistics 3.4 Consumption scenario in Europe 3.5 Use application summary 4. Environmental safety assessment 4.1 Environmental exposure assessment 4.1.1 Biotic and abiotic degradability 4.1.2 Removal 4.1.3 Monitoring studies 4.1.4 Exposure assessment: scenario description 4.1.5 Substance data used for the exposure calculation 4.1.6 PEC calculations 4.1.7 Bioconcentration 4.2 Environmental effects assessment 4.2.1 Ecotoxicity 4.2.1.1 Aquatic ecotoxicity 4.2.1.2 Terrestrial ecotoxicity 4.2.1.3 Sediment ecotoxicity 4.2.1.4 Ecotoxicity to sewage microorganisms 4.2.1.5 Reassurance on absence of estrogenic effects 4.2.2 PNEC calculations 4.2.2.1 Aquatic PNEC 4.2.2.2 Terrestrial PNEC 4.2.2.3 Sludge PNEC 4.2.2.4 Sediment PNEC 4.2.2.5 STP PNEC 4.3 Environment risk assessment 5. 5. References 6. Contributors to the report 6.1 Substance team 6.2 HERA environmental task force 6.3 HERA human health task force 6.4 Industry coalition for the OECD/ICCA SIDS assessment of LAS 2 2. Executive Summary Linear alkylbenzene sulphonate (LAS) is an anionic surfactant. It was introduced in 1964 as the readily biodegradable replacement for highly branched alkylbenzene sulphonates (ABS). -

Using Dynamic Combinatorial Chemistry to Construct Novel Thioester and Disulfide Lipids
Using dynamic combinatorial chemistry to construct novel thioester and disulfide lipids A dissertation submitted to the University of Manchester for the degree of MSc by Research Chemistry In the Faculty of Science and Engineering 2017 Jack A. Yung School of Chemistry Table of Contents List of figures, tables and equations 5 Symbols and abbreviations 10 Abstract 12 Declaration 13 Copyright statement 13 Acknowledgements 14 The author 14 Chapter 1. Introduction 15 1.1 The cell membrane 16 1.1.1 Function and composition 16 1.2 Membrane lipids 16 1.2.1 Lipid classification 16 1.2.2 Amphiphiles 17 1.2.3 Glycolipids 17 1.2.4 Sterols 18 1.2.5 Phospholipids 19 1.3 Lipid vesicles 20 1.3.1 Supramolecular self-assembly 20 1.3.2 Interaction free energies 21 1.3.3 Framework for the theory of self-assembly 22 1.3.4 Micelles 23 1.3.5 Lipid bilayers 24 1.3.6 Vesicles 25 1.3.7 Phase-transition temperature 27 1.4 Amphiphilic building blocks 27 1.5 Thioesters 29 1.5.1 Thioester reactivity 29 1.5.2 Trans-thioesterification 30 1.5.3 Thioester exchange reactions in DCC 31 1.6 Disulfides 32 2 1.6.1 Disulfide reactivity 32 1.6.2 Thiol-disulfide interchange reactions 32 1.6.3 Disulfide exchange reactions in DCC 33 1.7 Pre-biotic lipids 34 1.7.1 Sources of pre-biotic organic compounds 34 1.7.2 The first pre-biotic membrane structure 36 1.7.3 The role of sulfur in pre-biotic chemistry 36 1.8 Artificially designed vesicles 37 1.8.1 Applications of artificially designed vesicles 37 1.8.2 Zeta-potential 37 1.8.3 Vesicle design 38 1.9 Targets 39 1.9.1 Aims 39 Chapter 2. -

Variations in the Structure and Reactivity of Thioester Functionalized Self-Assembled Monolayers and Their Use for Controlled Surface Modification
Variations in the structure and reactivity of thioester functionalized self-assembled monolayers and their use for controlled surface modification Inbal Aped, Yacov Mazuz and Chaim N. Sukenik* Full Research Paper Open Access Address: Beilstein J. Nanotechnol. 2012, 3, 213–220. Department of Chemistry and Institute for Nanotechnology and doi:10.3762/bjnano.3.24 Advanced Materials, Bar-Ilan University, Ramat-Gan, Israel 52900 Received: 01 December 2011 Email: Accepted: 10 February 2012 Chaim N. Sukenik* - [email protected] Published: 09 March 2012 * Corresponding author This article is part of the Thematic Series "Self-assembly at solid surfaces". Keywords: siloxane-anchored self-assembled monolayers; sulfonated interfaces; Guest Editors: S. R. Cohen and J. Sagiv surface chemistry © 2012 Aped et al; licensee Beilstein-Institut. License and terms: see end of document. Abstract Thioester-functionalized, siloxane-anchored, self-assembled monolayers provide a powerful tool for controlling the chemical and physical properties of surfaces. The thioester moiety is relatively stable to long-term storage and its structure can be systematically varied so as to provide a well-defined range of reactivity and wetting properties. The oxidation of thioesters with different-chain- length acyl groups allows for very hydrophobic surfaces to be transformed into very hydrophilic, sulfonic acid-bearing, surfaces. Systematic variation in the length of the polymethylene chain has also allowed us to examine how imbedding reaction sites at various depths in a densely packed monolayer changes their reactivity. π-Systems (benzene and thiophene) conjugated to the thioester carbonyl enable the facile creation of photoreactive surfaces that are able to use light of different wavelengths. -

Chapter 7 - Alkenes and Alkynes I
Andrew Rosen Chapter 7 - Alkenes and Alkynes I 7.1 - Introduction - The simplest member of the alkenes has the common name of ethylene while the simplest member of the alkyne family has the common name of acetylene 7.2 - The (E)-(Z) System for Designating Alkene Diastereomers - To determine E or Z, look at the two groups attached to one carbon atom of the double bond and decide which has higher priority. Then, repeat this at the other carbon atom. This system is not used for cycloalkenes - If the two groups of higher priority are on the same side of the double bond, the alkene is designated Z. If the two groups of higher priority are on opposite sides of the double bond, the alkene is designated E - Hydrogenation is a syn/cis addition 7.3 - Relative Stabilities of Alkenes - The trans isomer is generally more stable than the cis isomer due to electron repulsions - The addition of hydrogen to an alkene, hydrogenation, is exothermic (heat of hydrogenation) - The greater number of attached alkyl groups, the greater the stability of an alkene 7.5 - Synthesis of Alkenes via Elimination Reactions, 7.6 - Dehydrohalogenation of Alkyl Halides How to Favor E2: - Reaction conditions that favor elimination by E1 should be avoided due to the highly competitive SN 1 mechanism - To favor E2, a secondary or tertiary alkyl halide should be used - If there is only a possibility for a primary alkyl halide, use a bulky base - Use a higher concentration of a strong and nonpolarizable base, like an alkoxide - EtONa=EtOH favors the more substituted double bond -
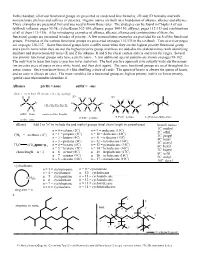
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

215-216 HH W12-Notes-Ch 15
Chem 215 F12 Notes Notes – Dr. Masato Koreeda - Page 1 of 17. Date: October 5, 2012 Chapter 15: Carboxylic Acids and Their Derivatives and 21.3 B, C/21.5 A “Acyl-Transfer Reactions” I. Introduction Examples: note: R could be "H" R Z R O H R O R' ester O carboxylic acid O O an acyl group bonded to R X R S acid halide* R' an electronegative atom (Z) thioester O X = halogen O R' R, R', R": alkyl, alkenyl, alkynyl, R O R' R N or aryl group R" amide O O O acid anhydride one of or both of R' and R" * acid halides could be "H" R F R Cl R Br R I O O O O acid fluoride acid chloride acid bromide acid iodide R Z sp2 hybridized; trigonal planar making it relatively "uncrowded" O The electronegative O atom polarizes the C=O group, making the C=O carbon "electrophilic." Resonance contribution by Z δ * R Z R Z R Z R Z C C C C O O O δ O hybrid structure The basicity and size of Z determine how much this resonance structure contributes to the hybrid. * The more basic Z is, the more it donates its electron pair, and the more resonance structure * contributes to the hybrid. similar basicity O R' Cl OH OR' NR'R" Trends in basicity: O weakest increasing basiciy strongest base base Check the pKa values of the conjugate acids of these bases. Chem 215 F12 Notes Notes –Dr. Masato Koreeda - Page 2 of 17.