(Silurian) Conodont Biostratigraphy of Saaremaa, Estonia, and a Correlation with Britain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rein Einasto
Rein Einasto Nähtust ja tehtust RAAMATU VÄLJAANDMIST ON TOETANUD Tõrva Kirik-Kammersaal, juhataja Ilmar Kõverik Pärtli Paas OÜ, juhataja Hillar Müür Lossikivi OÜ, juhataja Aivar Allikmaa Paekivitoodete Tehase OÜ, juhatuse esimees Vladimir Libman Väo Paas OÜ, juhataja Veljo Haube Reval Stone OÜ, juhataja Hindrek Auväärt Gildemann Dolomite Designs, Madis ja Mihkel Villand J. Viru Markšeideribüroo OÜ, IT-tugiisik, Priit Tammik Steiger Inseneribüroo OÜ, juhataja Erki Niitlaan Talter AS, mäetöödejuht Heini Viilup Redoil OÜ, juhataja Raivo Reinu Tunne Kelam, aatesõber Ülo Vooglaid, aatesõber Toimetanud Mari-Leen Toome Kujundanud Henry Vürst / Kujunduskuur Raamatus kasutatud fotod pärinevad autori erakogust Tallinna Tehnikakõrgkool, 2009 Pärnu mnt 62, 10135 Tallinn ISBN 978-9985-9906-8-1 KIVIS ON ELU JA MÕTISKELU Küllap Sa isegi tead, palju kivis on head. Palju kultuuri on kivis pikkade ajastute rivis. palju kivi – kultuuris ajaloosündmusis suuris. Kivis peidus on aegadetagune elu ja juurdleva vaimu mõtiskelu; kivi peidab inimelu võlu ja valu – hingetuid kivi kuuldavasti ei talu. Kivisse kantuna olevik põlistub, üleisikuline inimeses õilistub, kivis on paljude põlvkondade vaev, allaheidetute põlgus ja raev. Rahva elupõline maa-ootus ja kustumatu priiuse lootus. Vaadates kivile kestvamalt silma, tajume tagamaid, igavikulist ilma; püsiväärtused saavad puhtama värvi, argipäev ei ulatu rikkuma närvi... Rein Einasto Paevana SISUKORD EESSÕNA ....................................................................................................... 6 REIN -

Politsei Tabas Kuressaares Korteripeolt 15 Purjus Last Üks Nooruk Ähvardas Marientali De Patrullidega, Kuna Maja Seestpoolt Lukku,” Rääkis Sikk Su Alkoholi
“Ükskord Riigigümnaasium lindilõikamise ootel LK 2 • Kauneimad kodud: suur fotogalerii LK 6-8 nägin rada ainult läbi Taasiseseisvumis- prillidel päeval heiskame olevate riigilipu! väikeste pilude.” KUUEAASTANE Järgmine Saarte Hääl MOTOSPORTLANE ilmub laupäeval, 21.08 NIKLAS JOHANSON LK 4 Neljapäev, 19. august 2021 • Nr 135 (5389) • Hind 1,20 € VÄLJAKUTSELE KÕIK VABAD JÕUD: Politseil oli alust arvata, et tegemist tuleb suure hulga alkoholi tarvitanud noortega. Nii oligi. FOTOMONTAAŽ / LIINA ÕUN Politsei tabas Kuressaares korteripeolt 15 purjus last Üks nooruk ähvardas Marientali de patrullidega, kuna maja seestpoolt lukku,” rääkis Sikk su alkoholi. Aga oli ka neid, kes olid räägitud. Lugu ise oli Siku sõnul vägagi korterelamu kolmandal korrusel ukse ees oli kümmekond Marientali operatsioonist. jõudnud ära juua pudeli või rohkem.” õpetlik. Esmaspäeva pealelõuna ja õh- peetud peol aknast alla hüpata. jalgratast. Siiski õnnestus politseil Leiti ka sigarette ja e-sigaret. Siku tupoolik tunduvad olevat üsna süütu “Tuppa astudes oli tunda tugevat “Et oleks nende lastega ust lõhkumata korterisse sõnul võib arvata, et üht-teist jõudsid aeg, mil poleks nagu erilist põhjust las- alkoholilõhna,” ütleb politsei. seal jõudu tegeleda,” põh- pääseda ning üsna pea noored ka tualetipotist alla lasta. te tegemisi kontrollida, midagi taolist jendas Sikk, viidates, et oli saabus kohale ka korteri Tabatud noored viidi politseijaos- eeldaks pigem nädalavahetuselt. alust arvata, et tegemist tu- omanik. Pidutsejate vanus konda, koostati protokollid ning lapse- Tõenäoliselt mängis korterisse ko- Kadri Häng-Nuum, Raul Vinni leb suure hulga alkoholi tar- jäi vahemikku 13–17, ena- vanemad tulid võsukestele järele. gunemisel oma rolli vihmane ilm. [email protected] vitanud noortega. masti oli tegu 13–15-aastaste- Pealtnägijate sõnul käisid lapsevane- “Muidu oleks nad võib-olla kuskil Esmalt tabati trepikojast kaks ga. -

International Geological Correlation Programme. Project 591 "The Early to Middle Paleozoic Revolution"
International Geological Correlation Programme. Project 591 "The Early to Middle Paleozoic Revolution". Proceedings of the 3rd IGCP 591 Annual Meeting : Lund, Sweden, 9–19 June 2013 Lindskog, Anders; Mehlqvist, Kristina 2013 Link to publication Citation for published version (APA): Lindskog, A., & Mehlqvist, K. (Eds.) (2013). International Geological Correlation Programme. Project 591 "The Early to Middle Paleozoic Revolution". Proceedings of the 3rd IGCP 591 Annual Meeting : Lund, Sweden, 9–19 June 2013. (International Geological Correlation Programme, UNESCO; Vol. 3). Department of Geology, Lund University. Total number of authors: 2 General rights Unless other specific re-use rights are stated the following general rights apply: Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal Read more about Creative commons licenses: https://creativecommons.org/licenses/ Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. LUND UNIVERSITY PO Box 117 221 00 Lund +46 46-222 00 00 ANDERS LINDSKOG | KRISTINA MEHLQVIST Printed by Media-Tryck, Lund 2013 Proceedings of the 3rd IGCP 591 Annual Meeting Proceedings of the 3 Lund, Sweden, 9–19 June 2013 EDITED BY ANDERS LINDSKOG | KRISTINA MEHLQVIST DEPARTMENT OF GEOLOGY | LUND UNIVERSITY The abstracts within this volume were presented at the 3rd IGCP 591 Annual rd Meeting, which was held in Lund, Sweden, in June 2013. -

Lääne-Saare Valla Arengukava 2015–2023 (2030)
Lisa 1 Lääne-Saare vallavolikogu 21.08.2017 määruse nr 10 juurde LÄÄNE-SAARE VALLA ARENGUKAVA 2015–2023 (2030) Lääne-Saare valla arengukava aastateks 2015-2023(2030) Sisukord 1. LÄÄNE-SAARE VALLA PRAEGUNE OLUKORD JA ARENGUSUUNDUMUSED ......................... 4 1.1 ASUKOHT JA TERRITOORIUM ................................................................................................. 4 1.2 ASUSTUS JA RAHVASTIK ........................................................................................................... 6 1.3 TÖÖHÕIVE ................................................................................................................................... 9 2. KOHALIKU OMAVALITSUSE JUHTIMINE JA HALDAMINE ........................................................... 11 2.1. VALLA JUHTIMINE .............................................................................................................. 11 2.2. KOHALIK EELARVE ............................................................................................................. 11 2.3 KOONDHINNANG PRAEGUSELE OLUKORRALE ......................................................... 12 2.4 VALITSEMISEKS VAJALIKUD HOONED JA RAJATISED ...................................................... 13 3. LÄÄNE-SAARE VALLA ARENGUNÄGEMUS ....................................................................................... 14 3.1. VISIOON AASTAKS 2023 ........................................................................................................ 14 3.2. STRATEEGILISED EESMÄRGID ............................................................................................ -
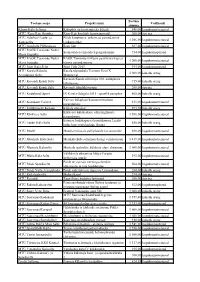
Toetuse Saaja Projekti Nimi Toetuse Summa Valdkond Kõnnu Küla
Toetuse Toetuse saaja Projekti nimi Valdkond summa Kõnnu Küla Seltsing Külaplats on kasutamiseks kõigile 848,00 kogukonnateenused MTÜ Karu-Kati Õpituba Karu-Kati koolitab lapsevanemaid 200,00 õpiring MTÜ Aderkasi Laulu- ja Pöide kaupluse e. seltsimaja parendamine 1 306,00 kogukonnateenused Tantsuselts (jätkuprojekt) MTÜ Anseküla Põllumeeste Eesti lipp 387,00 kogukonnateenused MTÜ EAÕK Tornimäe Neitsi Suunaviida ja infotahvli paigaldamine 734,00 kogukonnapärand Maria kogudus MTÜ EAÕK Tornimäe Neitsi EAÕK Tornimäe kirkuaia peavärava trepi ja 1 200,00 kogukonnateenused Maria kogudus värava parendamione MTÜ Jööri Küla Selts Jööri Folk 2013 394,00 kogukonnapärand MTÜ Karala Külaelu Karala sõprusküla Tremont Eesti X 1 008,00 kohalik areng Arendamise Selts Maapäeval Kavandi Kandi seltsimaja 100. aastapäeva MTÜ Kavandi Kandi Selts 715,00 kohalik areng tähistamine MTÜ Kavandi Kandi Selts Kavandi filmilektoorium 200,00 õpiring MTÜ Kodukandi Sport IX Karala Odagala 2013 - sportlik perepäev 860,00 kohalik areng Vaivere külaplatsi kasutusvõimaluste MTÜ Kodukant Vaivere 831,00 kogukonnateenused parandamine MTÜ Kultuuriselts Kevade Kõljala õunalaat 293,76 kohalik areng Kõrkvere külakeskuse olmetingimuste MTÜ Kõrkvere Selts 1 596,00 kogukonnateenused parandamine Esimese kadakapäeva korraldamine Leedri MTÜ Leedri Küla Selts 850,00 kohalik areng külas koos merelaulude õhtuga MTÜ Medel Meedla mõisaaia palliplatside korrastamine 888,00 kogukonnateenused MTÜ Metsküla Küla Selts Metsküla küla seltsimaja köögi ventilatsioon 1 439,00 kogukonnateenused MTÜ Mustjala -

Regionaaltoetused Saaremaal 2016
Regionaaltoetused Saaremaal 2016 Rahandusministeeriumi regionaalvaldkonna infoleht Infolehes on antud ülevaade 2016. aastal Saare maakonnas Rahandusministeeriumi rakendatavatest regionaalarengu programmidest toetust saanud projektidest. Rahandusministeeriumis on regionaalarengu toetamiseks ellu kutsutud ligi veerandsada toetusmeedet ja -programmi. 2016. aastal tehti toetusotsuseid enam kui 96 miljoni euro ulatuses, millele lisanduvad üle-Eestilised Euroopa territoriaalse koostöö programmide projektid 32 miljoni euroga. Saaremaal elluviidavate projektide toetuseks eraldati 2016. aastal 6,6 miljonit eurot. 1 Euroopa struktuuri- ja investeerimisfondide toetused 2016. aastal toetust saanud projektid: Toetuse saaja Projekti nimi Toetuse summa Piirkondlikud algatused tööhõive ja ettevõtlikkuse edendamiseks SA Saaremaa Saaremaa piirkondlike algatuste tugiprogramm 2015- 91 449 Arenduskeskus 2016 Piirkondade konkurentsivõime tugevdamise investeeringud Kuressaare Linnavalitsus Kuressaare linnakeskuse avaliku ruumi kaasajastamine 2 805 387 Kuressaare kogupere- ja elamuskeskus Hõbevalge (Thule OÜ Hõbekeskus koda) 2 011 202 Regionaalsete kompetentsikeskuste arendamine Tallinna Tehnikaülikool Väikelaevaehituse kompetentsikeskus 699 999 2016. aastal eraldati regionaalprojektide elluviimiseks Saaremaal kokku 5 608 039 eurot Euroopa struktuuri- ja investeerimisfondide vahendeid. Euroopa territoriaalse koostöö programmid Euroopa territoriaalse koostöö programmidest toetatakse projekte, mis suurendavad Euroopa äärealade, linnade, piirkondade ja regioonide -

AIP Estonia AD 2.EEKE-1 08 NOV 2018
eAIP Estonia AD 2.EEKE-1 08 NOV 2018 EEKE — Kuressaare EEKE AD 2.1 LENNUVÄLJA EEKE AD 2.1 AERODROME LOCATION ASUKOHAINDEKS JA NIMI INDICATOR AND NAME EEKE — Kuressaare EEKE AD 2.2 LENNUVÄLJA EEKE AD 2.2 AERODROME GEO- GEOGRAAFILISED JA GRAPHICAL AND ADMINISTRATIVE ADMINISTRATIIVANDMED DATA ARP koordinaadid ja asukoht 581348N 0223034E 1 lennuväljal 1237 m / 173° MAG lävest 17, radade ristumiskohas. ARP coordinates and site at AD 1237 m / 173° MAG FM THR 17, on the INT of RWYs. Suund ja kaugus Kuressaarest 2 Direction and distance from Kures- 1.5 NM SSE FM Kuressaare saare Kõrgus merepinnast / Keskmine 3 kõrgeim temperatuur 15 ft / 21.0 °C Elevation / Reference temperature Geoidi hälve lennuväljal 4 66 ft Geoid undulation at AD MAG VAR / Aastane muutus 5 7° E (2015) / +0.2° MAG VAR / Annual change Lennuvälja haldaja, aadress, telefon, Aadress: AS Tallinna Lennujaam Kuressaare lennujaam telefaks, e-mail, AFS, URL Roomassaare 1 AD operator, address, telephone, 93815 Kuressaare telefax, e-mail, AFS, URL ESTONIA 6 Tel: +372 45 30 313 Faks: +372 45 30 340 E-post: [email protected] AFS: EEKEZTZX URL: www.kuressaare-airport.ee Lubatud liikluse liigid (IFR/VFR) 7 IFR/VFR Types of traffic permitted (IFR/VFR) Märkused Ei ole 8 Remarks NIL EEKE AD 2.3 TÖÖAJAD EEKE AD 2.3 OPERATIONAL HOURS Lennuvälja haldaja MON-FRI : 0700-1500 (0600-1400) AD operator 1 Lennuvälja tööaeg Sama mis p 8. AD Operational hours Same as p 8. Toll ja migratsioon 2 MON-SUN: 0600-2000 (0500-1900) Customs and immigration Tervishoid MON-FRI: 0600-1600 (0500-1500); 3 Health and sanitation SAT-SUN: 0600-1500 (0500-1400) AIS Briifing Olemas self-briifingu kasutamisvõimalus või teenust osutab ATS. -

Detroit River Group in the Michigan Basin
GEOLOGICAL SURVEY CIRCULAR 133 September 1951 DETROIT RIVER GROUP IN THE MICHIGAN BASIN By Kenneth K. Landes UNITED STATES DEPARTMENT OF THE INTERIOR Oscar L. Chapman, Secretary GEOLOGICAL SURVEY W. E. Wrather, Director Washington, D. C. Free on application to the Geological Survey, Washington 25, D. C. CONTENTS Page Page Introduction............................ ^ Amherstburg formation................. 7 Nomenclature of the Detroit River Structural geology...................... 14 group................................ i Geologic history ....................... ^4 Detroit River group..................... 3 Economic geology...................... 19 Lucas formation....................... 3 Reference cited........................ 21 ILLUSTRATIONS Figure 1. Location of wells and cross sections used in the study .......................... ii 2. Correlation chart . ..................................... 2 3. Cross sections A-«kf to 3-G1 inclusive . ......................;.............. 4 4. Facies map of basal part of Dundee formation. ................................. 5 5. Aggregate thickness of salt beds in the Lucas formation. ........................ 8 6. Thickness map of Lucas formation. ........................................... 10 7. Thickness map of Amherstburg formation (including Sylvania sandstone member. 11 8. Lime stone/dolomite facies map of Amherstburg formation ...................... 13 9. Thickness of Sylvania sandstone member of Amherstburg formation.............. 15 10. Boundary of the Bois Blanc formation in southwestern Michigan. -

Koolieelsete Lasteasutuste Teeninduspiirkondade Kinnitamine“ LISA 1
Kihelkonna Vallavolikogu määruse nr 14 „Koolieelsete lasteasutuste teeninduspiirkondade kinnitamine“ LISA 1 Saaremaa ühinevate omavalitsuste ametiasutuste hallatavate koolieelsete lasteasutuste teeninduspiirkonnad 1. Aste Lasteaia teeninduspiirkond on Anijala, Ansi, Aste, Asuküla, Aula-Vintri, Eikla, Endla, Haamse, Hakjala, Hübja, Irase, Jootme, Jõe, Kaarma, Kaarma-Kirikuküla, Kaarmise, Kaisvere, Kaubi, Kellamäe, Keskvere, Kiratsi, Koidu, Koidula, Kuke, Kungla, Käku, Laadjala, Laoküla, Maleva, Meedla, Metsaküla, Mullutu, Nõmme, Piila, Põlluküla, Pähkla, Pärni, Randvere, Saia, Sepa, Tamsalu, Tõlli, Tõrise, Tõru, Uduvere, Unimäe, Vantri, Vestla, Viira, Õha külad ning Aste alevik. 2. Kaali Kooli teeninduspiirkond on Eiste, Ennu, Haeska, Hämmelepa, Iilaste, Ilpla, Kaali, Kailuka, Kangrusselja, Kiritu, Kuusiku, Kõljala, Kõnnu, Laheküla, Leina, Liiva, Liiva- Putla, Masa, Matsiranna, Metsaküla, Mustla, Nässuma, Pihtla, Püha, Rahniku, Rannaküla, Reeküla, Reo, Räimaste, Sagariste, Salavere, Sandla, Sauaru, Saue-Putla, Sepa, Sutu, Suure-Rootsi, Tõlluste, Vanamõisa, Väike-Rootsi, Väljaküla külad. 3. Kahtla Lasteaed-Põhikooli teeninduspiirkond on Aaviku, Asva, Audla, Jõe, Kahtla, Kapra, Kingli, Kõiguste, Käo, Laheküla, Laimjala, Mustla, Mägi-Kurdla, Nõmme, Pahavalla, Paju-Kurdla, Randvere, Rannaküla, Ridala, Ruhve, Saareküla, Saaremetsa, Viltina, Üüvere külad. 4. Kihelkonna Kooli teeninduspiirkond on Abaja, Abula, Kallaste, Kalmu, Karujärve, Kehila, Kiirassaare, Kotsma, Kuralase, Kuremetsa, Kurevere, Kuumi, Kuusiku, Kõruse, Kõõru, Liiva, Loona, Lätiniidi, -

Xerox University Microfilms
information t o u s e r s This material was produced from a microfilm copy of the original document. While the most advanced technological means to photograph and reproduce this document have been used, the quality is heavily dependent upon the quality of the original submitted. The following explanation of techniques is provided to help you understand markings or patterns which may appear on this reproduction. 1.The sign or "target” for pages apparently lacking from the document photographed is "Missing Page(s)". If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting thru an image and duplicating adjacent pages to insure you complete continuity. 2. When an image on the film is obliterated with a large round black mark, it is an indication that the photographer suspected that the copy may have moved during exposure and thus cause a blurred image. You will find a good image of the page in the adjacent frame. 3. When a map, drawing or chart, etc., was part of the material being photographed the photographer followed a definite method in "sectioning" the material. It is customary to begin photoing at the upper left hand corner of a large sheet and to continue photoing from left to right in equal sections with a small overlap. If necessary, sectioning is continued again - beginning below the first row and continuing on until complete. 4. The majority of usefs indicate that the textual content is of greatest value, however, a somewhat higher quality reproduction could be made from "photographs" if essential to the understanding of the dissertation. -

Saaremaa Vald Külad 1) Aaviku 2) Abaja 3) Abruka 4) Abula 5
Saaremaa vald Külad 1) Aaviku 50) Jaani 99) Kaunispe 2) Abaja 51) Jauni 100) Kavandi 3) Abruka 52) Jootme 101) Kehila 4) Abula 53) Jursi 102) Kellamäe 5) Allikalahe 54) Jõe 103) Keskranna 6) Anepesa 55) Jõelepa 104) Keskvere 7) Angla 56) Jõempa 105) Kihelkonna-Liiva 8) Anijala 57) Jõgela 106) Kiirassaare 9) Anseküla 58) Jõiste 107) Kingli 10) Ansi 59) Jämaja 108) Kipi 11) Arandi 60) Järise 109) Kiratsi 12) Ardla 61) Järve 110) Kirderanna 13) Are 62) Järveküla 111) Kiritu 14) Ariste 63) Jööri 112) Kiruma 15) Arju 64) Kaali 113) Kogula 16) Aru 65) Kaali-Liiva 114) Koidula 17) Aruste 66) Kaarma 115) Koiduvälja 18) Aste 67) Kaarma-Jõe 116) Koigi 19) Asuka 68) Kaarma-Kirikuküla 117) Koigi-Väljaküla 20) Asuküla 69) Kaarma-Kungla 118) Koikla 21) Asva 70) Kaarmise 119) Koimla 22) Atla 71) Kaavi 120) Koki 23) Audla 72) Kahtla 121) Koksi 24) Aula-Vintri 73) Kahutsi 122) Koovi 25) Austla 74) Kailuka 123) Kopli 26) Easte 75) Kaimri 124) Kotlandi 27) Eeriksaare 76) Kaisa 125) Kotsma 28) Eikla 77) Kaisvere 126) Kugalepa 29) Eiste 78) Kakuna 127) Kuiste 30) Endla 79) Kalju 128) Kuke 31) Ennu 80) Kallaste 129) Kungla 32) Haamse 81) Kallemäe 130) Kuninguste 33) Haapsu 82) Kalli 131) Kuralase 34) Haeska 83) Kalma 132) Kuremetsa 35) Hakjala 84) Kalmu 133) Kurevere 36) Hiievälja 85) Kandla 134) Kuumi 37) Himmiste 86) Kangrusselja 135) Kuuse 38) Hindu 87) Kanissaare 136) Kuusiku 39) Hirmuste 88) Kapra 137) Kuusnõmme 40) Hämmelepa 89) Karala 138) Kõiguste 41) Hänga 90) Kareda 139) Kõinastu 42) Hübja 91) Kargi 140) Kõljala 43) Iide 92) Karida 141) Kõnnu 44) -

BIASES in INTERPRETATION of the FOSSIL RECORD of CONODONTS by MARK A
[Special Papers in Palaeontology, 73, 2005, pp. 7–25] BETWEEN DEATH AND DATA: BIASES IN INTERPRETATION OF THE FOSSIL RECORD OF CONODONTS by MARK A. PURNELL* and PHILIP C. J. DONOGHUE *Department of Geology, University of Leicester, University Road, Leicester LE1 7RH, UK; e-mail: [email protected] Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queens Road, Bristol BS8 1RJ, UK; e-mail: [email protected] Abstract: The fossil record of conodonts may be among and standing generic diversity. Analysis of epoch ⁄ stage-level the best of any group of organisms, but it is biased nonethe- data for the Ordovician–Devonian interval suggests that less. Pre- and syndepositional biases, including predation there is generally no correspondence between research effort and scavenging of carcasses, current activity, reworking and and generic diversity, and more research is required to bioturbation, cause loss, redistribution and breakage of ele- determine whether this indicates that sampling of the cono- ments. These biases may be exacerbated by the way in which dont record has reached a level of maturity where few genera rocks are collected and treated in the laboratory to extract remain to be discovered. One area of long-standing interest elements. As is the case for all fossils, intervals for which in potential biases and the conodont record concerns the there is no rock record cause inevitable gaps in the strati- pattern of recovery of different components of the skeleton graphic distribution of conodonts, and unpreserved environ- through time. We have found no evidence that the increas- ments lead to further impoverishment of the recorded ing abundance of P elements relative to S and M elements spatial and temporal distributions of taxa.