The Biogeography and Community Structure of Kelp Forest Macroinvertebrates Laurel A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Thesis Presented to the Faculty of the Department of Marine Science
COMMUNITY STRUCTURE AND ENERGY FLOW WITHIN RHODOLITH HABITATS AT SANTA CATALINA ISLAND, CA A Thesis Presented to The Faculty of the Department of Marine Science San José State University In Partial Fulfillment Of the Requirements for the Degree Master of Science In Marine Science by Scott Stanley Gabara December 2014 © 2014 Scott S. Gabara ALL RIGHTS RESERVED The Designated Thesis Committee Approves the Thesis Titled COMMUNITY STRUCTURE AND ENERGY FLOW WITHIN RHODOLITH HABITATS AT SANTA CATALINA ISLAND, CA By Scott Stanley Gabara APPROVED FOR THE DEPARTMENT OF MARINE SCIENCE SAN JOSÉ STATE UNIVERSITY December 2014 Dr. Diana L. Steller Moss Landing Marine Laboratories Dr. Michael H. Graham Moss Landing Marine Laboratories Dr. Scott L. Hamilton Moss Landing Marine Laboratories ABSTRACT COMMUNITY STRUCTURE AND ENERGY FLOW WITHIN RHODOLITH HABITATS AT SANTA CATALINA ISLAND, CA by Scott Stanley Gabara The purpose of this study was to describe the floral and faunal community associated with rhodolith beds, which are aggregations of free-living coralline algal nodules, off of Santa Catalina Island. Surveys of macroalgal cover, infaunal and epifaunal invertebrates, and fishes suggest rhodolith beds off Santa Catalina Island support greater floral and faunal abundances than adjacent sand habitat. Community separation between rhodolith and sand habitats was due to increased presence of fleshy macroalgae, herbivorous gastropods, and greater abundance of infaunal invertebrates dominated by amphipods, mainly tanaids and gammarids. Stable isotopes were used to determine important sources of primary production supporting rhodolith beds and to identify the major pathways of energy. Stable isotopes suggest the rhodolith bed food web is detrital based with contributions from water column particulate organic matter, drift kelp tissue, and kelp particulates from adjacent kelp beds. -

OREGON ESTUARINE INVERTEBRATES an Illustrated Guide to the Common and Important Invertebrate Animals
OREGON ESTUARINE INVERTEBRATES An Illustrated Guide to the Common and Important Invertebrate Animals By Paul Rudy, Jr. Lynn Hay Rudy Oregon Institute of Marine Biology University of Oregon Charleston, Oregon 97420 Contract No. 79-111 Project Officer Jay F. Watson U.S. Fish and Wildlife Service 500 N.E. Multnomah Street Portland, Oregon 97232 Performed for National Coastal Ecosystems Team Office of Biological Services Fish and Wildlife Service U.S. Department of Interior Washington, D.C. 20240 Table of Contents Introduction CNIDARIA Hydrozoa Aequorea aequorea ................................................................ 6 Obelia longissima .................................................................. 8 Polyorchis penicillatus 10 Tubularia crocea ................................................................. 12 Anthozoa Anthopleura artemisia ................................. 14 Anthopleura elegantissima .................................................. 16 Haliplanella luciae .................................................................. 18 Nematostella vectensis ......................................................... 20 Metridium senile .................................................................... 22 NEMERTEA Amphiporus imparispinosus ................................................ 24 Carinoma mutabilis ................................................................ 26 Cerebratulus californiensis .................................................. 28 Lineus ruber ......................................................................... -

The Diet and Predator-Prey Relationships of the Sea Star Pycnopodia Helianthoides (Brandt) from a Central California Kelp Forest
THE DIET AND PREDATOR-PREY RELATIONSHIPS OF THE SEA STAR PYCNOPODIA HELIANTHOIDES (BRANDT) FROM A CENTRAL CALIFORNIA KELP FOREST A Thesis Presented to The Faculty of Moss Landing Marine Laboratories San Jose State University In Partial Fulfillment of the Requirements for the Degree Master of Arts by Timothy John Herrlinger December 1983 TABLE OF CONTENTS Acknowledgments iv Abstract vi List of Tables viii List of Figures ix INTRODUCTION 1 MATERIALS AND METHODS Site Description 4 Diet 5 Prey Densities and Defensive Responses 8 Prey-Size Selection 9 Prey Handling Times 9 Prey Adhesion 9 Tethering of Calliostoma ligatum 10 Microhabitat Distribution of Prey 12 OBSERVATIONS AND RESULTS Diet 14 Prey Densities 16 Prey Defensive Responses 17 Prey-Size Selection 18 Prey Handling Times 18 Prey Adhesion 19 Tethering of Calliostoma ligatum 19 Microhabitat Distribution of Prey 20 DISCUSSION Diet 21 Prey Densities 24 Prey Defensive Responses 25 Prey-Size Selection 27 Prey Handling Times 27 Prey Adhesion 28 Tethering of Calliostoma ligatum and Prey Refugia 29 Microhabitat Distribution of Prey 32 Chemoreception vs. a Chemotactile Response 36 Foraging Strategy 38 LITERATURE CITED 41 TABLES 48 FIGURES 56 iii ACKNOWLEDGMENTS My span at Moss Landing Marine Laboratories has been a wonderful experience. So many people have contributed in one way or another to the outcome. My diving buddies perse- vered through a lot and I cherish our camaraderie: Todd Anderson, Joel Thompson, Allan Fukuyama, Val Breda, John Heine, Mike Denega, Bruce Welden, Becky Herrlinger, Al Solonsky, Ellen Faurot, Gilbert Van Dykhuizen, Ralph Larson, Guy Hoelzer, Mickey Singer, and Jerry Kashiwada. Kevin Lohman and Richard Reaves spent many hours repairing com puter programs for me. -

Marine Ecology Progress Series 457:85
This authors' personal copy may not be publicly or systematically copied or distributed, or posted on the Open Web, except with written permission of the copyright holder(s). It may be distributed to interested individuals on request. Vol. 457: 85–99, 2012 MARINE ECOLOGY PROGRESS SERIES Published June 21 doi: 10.3354/meps09693 Mar Ecol Prog Ser Geographic variation in demography of a temperate reef snail: importance of multiple life-history traits Rebecca G. Martone1,2,*, Fiorenza Micheli1 1Hopkins Marine Station, Stanford University, 120 Oceanview Blvd., Pacific Grove, California 93950, USA 2Present address: Institute for Resources, Environment and Sustainability, The University of British Columbia, Aquatic Ecosystem Research Laboratory, 429-2202 Main Mall, Vancouver, British Columbia V6T 1Z4, Canada ABSTRACT: Individual- and population-level performance may reflect trade-offs between energy allocation to different key demographic processes, such as growth and reproduction, which can, in turn, be influenced by local biotic and abiotic conditions. We explored geographic variation in demographic rates of an exploited benthic species, the wavy-turban snail Megastraea undosa, along the Pacific coast of Baja California, Mexico. We compared key life-history traits (i.e. fecun- dity, size at maturity, growth, and survivorship) of populations existing between 20 and 170 km apart under different conditions of ocean temperature and food availability. Trade-offs between growth and reproduction were evident across this environmental gradient, with higher growth rates in warmer locations leading to lower size-specific investment in gonad production. Because later onset of reproduction in populations from warmer areas was compensated by greater fecun- dity at larger sizes, geographic variation in life-history strategies resulted in similar age-specific reproductive output among different populations. -

PISCO 'Mobile' Inverts 2017
PISCO ‘Mobile’ Inverts 2017 Lonhart/SIMoN MBNMS NOAA Patiria miniata (formerly Asterina miniata) Bat star, very abundant at many sites, highly variable in color and pattern. Typically has 5 rays, but can be found with more or less. Lonhart/SIMoN MBNMS NOAA Patiria miniata Bat star (formerly Asterina miniata) Lonhart/SIMoN MBNMS NOAA Juvenile Dermasterias imbricata Leather star Very smooth, five rays, mottled aboral surface Adult Dermasterias imbricata Leather star Very smooth, five rays, mottled aboral surface ©Lonhart Henricia spp. Blood stars Long, tapered rays, orange or red, patterned aboral surface looks like a series of overlapping ringlets. Usually 5 rays. Lonhart/SIMoN MBNMS NOAA Henricia spp. Blood star Long, tapered rays, orange or red, patterned aboral surface similar to ringlets. Usually 5 rays. (H. sanguinolenta?) Lonhart/SIMoN MBNMS NOAA Henricia spp. Blood star Long, tapered rays, orange or red, patterned aboral surface similar to ringlets. Usually 5 rays. Lonhart/SIMoN MBNMS NOAA Orthasterias koehleri Northern rainbow star Mottled red, orange and yellow, large, long thick rays Lonhart/SIMoN MBNMS NOAA Mediaster aequalis Orange star with five rays, large marginal plates, very flattened. Confused with Patiria miniata. Mediaster aequalis Orange star with five rays, large marginal plates, very flattened. Can be mistaken for Patiria miniata Pisaster brevispinus Short-spined star Large, pale pink in color, often on sand, thick rays Lonhart/SIMoN MBNMS NOAA Pisaster giganteus Giant-spined star Spines circled with blue ring, thick -

See Life Trunk
See Life Trunk CABRILLO NATIONAL MONUMENT Objective The See Life Trunk is designed to help students experience the vast biodiversity of earth’s marine ecosystems from the comfort of their classroom. The activities, books, and DVDs in this trunk specifically target third and fourth grade students based on Next Generation Science Standards. The trunk will bring to life the meaning of biodiversity in the ocean, its role in the maintenance and function of healthy marine ecosystems, and what students can do to help protect this environment for generations to come. What’s Inside Books: In One Tidepool: Crabs, Snails and Salty Tails SEASHORE (One Small Square series) CORAL REEFS (One Small Square series) The Secrets of Kelp Forests The Secrets of the Tide Pools Shells of San Diego DVDs: Eyewitness Life Eyewitness Seashore Eyewitness Ocean Bill Nye the Science Guy: Ocean Life On the Edge of Land and Sea Activities, Resources & Worksheets: Marine Bio-Bingo Guess Who: Intertidal Patterns in Nature See Life & Habitats o Classroom set of Michael Ready photographs o 3D-printed biomodels Science Sampler Intro to Nature Journaling o Creature Features o Baseball Cards Who Am I? Beyond the Classroom Activities Intertidal Exploration 3D Cabrillo Bioblitz Beach clean-up How to Use this Trunk The See Life Trunk is designed to be used in a variety of ways. Most of the activities in the trunk can be adapted for any number of people for any amount of time, but some activities are better suited for entire-class participation (i.e. watching DVDs, Nature Journaling) while others are better suited for small groups (i.e. -
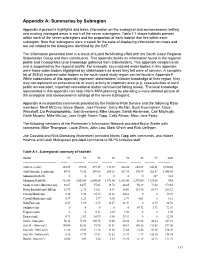
Appendix A: Summaries by Subregion
Appendix A: Summaries by Subregion Appendix A presents highlights and basic information on the ecological and socioeconomic setting and existing managed areas in each of the seven subregions. Table I-1 shows habitats present within each of the seven subregions and the proportion of each habitat that lies within each subregion. Note that subregions were created for the ease of displaying information on maps and are not related to the bioregions identified by the SAT. The information presented here is a result of a joint fact-finding effort with the South Coast Regional Stakeholder Group and their constituents. This appendix builds on information found in the regional profile and incorporates local knowledge gathered from stakeholders. This appendix complements and is supported by the regional profile. For example, key impaired water bodies in this appendix were those water bodies highlighted by stakeholders as areas they felt were of concern. A complete list of 303(d) impaired water bodies in the south coast study region can be found in Appendix F. While subsections of this appendix represent stakeholders’ intimate knowledge of their region, they may not represent an exhaustive list of every activity or important area (e.g. uses/activities at each public access point, important recreational and/or commercial fishing areas). The local knowledge represented in this appendix can help inform MPA planning by providing a more detailed picture of the ecological and socioeconomic settings of the seven subregions. Appendix A incorporates comments provided by the National Park Service and the following RSG members: Merit McCrea, Bruce Steele, Jack Peveler, Gerry Richter, Buck Everingham, Dave Weeshoff, Lia Protopapadakis, Joel Greenberg, Mike Gauger, Sarah Abramson, Carl Mayhugh, Garth Murphy, Mike McCoy, Jack Engle, Norris Tapp, Calla Allison, Mary Jane Foley. -

WSN Long Program 2014 FINAL
Western Society of Naturalists Meeting Program Tacoma, WA Nov. 13-16, 2014 1 Western Society of Naturalists Treasurer President ~ 2014 ~ Andrew Brooks Steven Morgan Dept of Ecology, Evolution Bodega Marine Laboratory, Website and Marine Biology UC Davis www.wsn-online.org UC Santa Barbara P.O. Box 247 Santa Barbara, CA 93106 Bodega, CA 94923 Secretariat [email protected] [email protected] Michael Graham Scott Hamilton Member-at-Large Diana Steller President-Elect Phil Levin Moss Landing Marine Laboratories Northwest Fisheries Science Gretchen Hofmann 8272 Moss Landing Rd Center Dept. Ecology, Evolution, & Moss Landing, CA 95039 Conservation Biology Division Marine Biology Seattle, WA 98112 Corey Garza UC Santa Barbara [email protected] CSU Monterey Bay Santa Barbara, CA 93106 [email protected] Seaside, CA 93955 [email protected] 95TH ANNUAL MEETING NOVEMBER 13-16, 2014 IN TACOMA, WASHINGTON Registration and Information Welcome! The registration desk will be open Thurs 1600-2000, Fri-Sat 0730-1800, and Sun 0800-1000. Registration packets will be available at the registration table for those members who have pre-registered. Those who have not pre-registered but wish to attend the meeting can pay for membership and registration (with a $20 late fee) at the registration table. Unfortunately, banquet tickets cannot be sold at the meeting because the hotel requires final counts of attendees well in advance. The Attitude Adjustment Hour (AAH) is included in the registration price, so you will only need to show your badge for admittance. WSN t-shirts and other merchandise can be purchased or picked up at the WSN Student Committee table. -

Sea Star Species Affected by Wasting Syndrome: (Updated 5/7/18)
Sea Star Species Affected by SSWS pacificrockyintertidal.org Last updated 2018-05-07 seastarwasting.org Sea Star Species Affected by Wasting Syndrome: (updated 5/7/18) High Mortality Solaster dawsoni (morning sun star) Evasterias troschelii (mottled star) Pisaster brevispinus (giant pink star) Pisaster ochraceus (ochre/purple star) Pycnopodia helianthoides (sunflower star) Some Mortality Patiria (Asterina) miniata (bat star) Dermasterias imbricata (leather star) Solaster stimpsoni (striped sun star) Orthasterias koehleri (rainbow star) Pisaster giganteus (giant star) Henricia spp. (blood star) Leptasterias spp (six-armed star) Luidia foliolata (sand star) Likely affected, mortality level not well documented Astropecten spp. (sand star) Mediaster aequalis (vermilion star) Linckia columbiae (fragile star) Pteraster tesselatus (slime star) Pteraster militaris (wrinkled star) Lophaster furcilliger vexator (crested star) Crossaster papposus (rose star) Astrometis sertulifera (fragile rainbow star) Stylasterias forreri (velcro star) Asterias amurensis (northern pacific star) ©2018 by University of California, Santa Cruz. All rights reserved. Page 1 of 2 Sea Star Species Affected by SSWS pacificrockyintertidal.org Last updated 2018-05-07 seastarwasting.org Order Paxillosida Family Astropectinidae Astropecten spp. (sand star) Family Luidiidae Luidia foliolata (sand star) Order Valvatida Family Asterinidae Patiria (Asterina) miniata (bat star) Family Goniasteridae Mediaster aequalis (vermilion star) Family Ophidiasteridae Linckia columbiae (fragile -

Interpreting the Paleozoogeography and Sea Level History of Thermally
Monographs of the Western North American Naturalist Volume 7 8th California Islands Symposium Article 6 9-9-2014 Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California Daniel R. Muhs U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Lindsey T. Groves Natural History Museum of Los Angeles County, [email protected] R. Randall Schumann U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Follow this and additional works at: https://scholarsarchive.byu.edu/mwnan Recommended Citation Muhs, Daniel R.; Groves, Lindsey T.; and Schumann, R. Randall (2014) "Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California," Monographs of the Western North American Naturalist: Vol. 7 , Article 6. Available at: https://scholarsarchive.byu.edu/mwnan/vol7/iss1/6 This Monograph is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Monographs of the Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Monographs of the Western North American Naturalist 7, © 2014, pp. 82–108 INTERPRETING THE PALEOZOOGEOGRAPHY AND SEA LEVEL HISTORY OF THERMALLY ANOMALOUS MARINE TERRACE FAUNAS: A CASE STUDY FROM THE LAST INTERGLACIAL COMPLEX OF SAN CLEMENTE ISLAND, CALIFORNIA Daniel R. Muhs1,3, Lindsey T. Groves2, and R. Randall Schumann1 ABSTRACT.—Marine invertebrate faunas with mixtures of extralimital southern and extralimital northern faunal elements, called thermally anomalous faunas, have been recognized for more than a century in the Quaternary marine terrace record of the Pacific Coast of North America. -
Nutritional Constraints on the Southern Sea Otter in the Monterey Bay National Marine Sanctuary
NUTRITIONAL CONSTRAINTS ON THE SOUTHERN SEA OTTER IN THE MONTEREY BAY NATIONAL MARINE SANCTUARY and a comparison to sea otter populations at San Nicolas Island, California and Glacier Bay, Alaska. Joint Final Report to Monterey Bay National Marine Sanctuary (and Monterey Bay Sanctuary Foundation) and the Marine Mammal Commission. Prepared by: Olav T. Oftedal PhD, Nutrition Laboratory, Conservation Ecology Center, Smithsonian’s National Zoological Park, Washington DC Katherine Ralls PhD, Center for Conservation and Evolutionary Genetics, Smithsonian’s National Zoological Park, Washington, DC M. Tim Tinker PhD, University of California Santa Cruz, Santa Cruz, California Alice Green, PhD candidate, Nutritional Biology, University of California Davis, Davis, California With assistance and contributions from: Seth Newsome PhD, Carnegie Institute of Washington, Washington, DC Jim Bodkin MS, Alaska Science Center, US Geological Survey, Anchorage, Alaska Gena Bentall MS, University of California Santa Cruz, Santa Cruz, California Nutritional constraints in the southern sea otter. Oftedal, Ralls, Tinker and Green, 2007 NUTRITIONAL CONSTRAINTS ON THE SOUTHERN SEA OTTER IN THE MONTEREY BAY NATIONAL MARINE SANCTUARY and a comparison to sea otter populations at San Nicolas Island, California and Glacier Bay, Alaska. O. Oftedal, K. Ralls, M.T. Tinker and A. Green with assistance and contributions from S. Newsome, J. Bodkin and G. Bentall Executive Summary The Monterey Bay National Marine Sanctuary in California, which extends from Marin in the north to Cambria in the south, encompasses much of the range of the threatened southern sea otter, Enhydra lutris nereis. The growth of this otter population has slowed and/or stalled due to increased mortality, and many otters die of disease. -
2020 Program
Western Society of Naturalists 101st Meeting Program Western Society of Naturalists ~ 2020 ~ Secretariat Chris Harley Patrick Martone Mary O’Connor Depts. Of Botany and Zoology University of British Columbia Vancouver, BC V6T1z4 Francis Janes UVIC [email protected] 101ST ANNUAL MEETING NOVEMBER 5 - NOVEMBER 8 2020 VIRTUAL MEETING 2 Welcome to the 101st Annual Meeting of the Western Society of Naturalists! WSN’s roots can be traced back to 1910 when a group of biologists, concerned about the lack of scientific meetings on the west coast, formed the Biological Society of the Pacific (call this our larval phase). The BSP was intended to include “any person interested in scientific work of a research nature.” In 1915, the American Society of Naturalists invited the BSP to join as a Pacific chapter under the ASN banner. Although the BSP voted in favor, the merger was eventually rejected by the ASN Executive Committee, who felt that we might not be sufficiently selective in admitting members. Through this process, the BSP formally metamorphosed into the Western Society of Naturalists in 1916. The first annual meeting of the society that would become WSN featured four scientific presentations and a dinner. Since then, our meetings and our society have grown substantially, but we still proudly welcome everyone who is interested in scientific work of a research nature. This year, we celebrate our 101st meeting. The circumstances are unusual, we are in the midst of a global pandemic, and also in a period of increased commitment to our society’s diversity, equity and inclusion of all naturalists.