A Thesis Presented to the Faculty of the Department of Marine Science
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Effects of Ocean Warming and Acidification on Fertilization Success and Early Larval Development in the Green Sea Urchin, Lytechinus Variegatus Brittney L
Nova Southeastern University NSUWorks HCNSO Student Theses and Dissertations HCNSO Student Work 12-1-2017 Effects of Ocean Warming and Acidification on Fertilization Success and Early Larval Development in the Green Sea Urchin, Lytechinus variegatus Brittney L. Lenz Nova Southeastern University, [email protected] Follow this and additional works at: https://nsuworks.nova.edu/occ_stuetd Part of the Marine Biology Commons, and the Oceanography and Atmospheric Sciences and Meteorology Commons Share Feedback About This Item NSUWorks Citation Brittney L. Lenz. 2017. Effects of Ocean Warming and Acidification on Fertilization Success and Early Larval Development in the Green Sea Urchin, Lytechinus variegatus. Master's thesis. Nova Southeastern University. Retrieved from NSUWorks, . (457) https://nsuworks.nova.edu/occ_stuetd/457. This Thesis is brought to you by the HCNSO Student Work at NSUWorks. It has been accepted for inclusion in HCNSO Student Theses and Dissertations by an authorized administrator of NSUWorks. For more information, please contact [email protected]. Thesis of Brittney L. Lenz Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science M.S. Marine Biology Nova Southeastern University Halmos College of Natural Sciences and Oceanography December 2017 Approved: Thesis Committee Major Professor: Joana Figueiredo Committee Member: Nicole Fogarty Committee Member: Charles Messing This thesis is available at NSUWorks: https://nsuworks.nova.edu/occ_stuetd/457 HALMOS COLLEGE OF NATURAL SCIENCES AND -

California State University, Northridge the Effects Of
CALIFORNIA STATE UNIVERSITY, NORTHRIDGE THE EFFECTS OF LECTINS IN SEA URCHIN LYTECHINUS PICTUS DURING GASTRULATION IN LOW CALCIUM SEA WATER A thesis submitted in partial fulfillment of the requirements For the degree of Master of Science in Biology By Siavash Nikkhou December 2013 The thesis of Siavash Nikkhou is approved by: ---------------------------------------------------- ----------------------------------------- Dr. Aida Metzenberg Date ---------------------------------------------------- ----------------------------------------- Dr. Stan Metzenberg Date ---------------------------------------------------- ----------------------------------------- Dr. Steven B. Oppenheimer, Chair Date California State University, Northridge ii Acknowledgements I would like to sincerely thank Dr. Steven B. Oppenheimer for believing in me and being the best mentor and an advisor a graduate can ask for and with his well rounded knowledge in the field assisted me throughout the research. I would like to thank the entire Biology faculty more specifically I would like to thank Dr Karels, Dr. Aida Metzenberg and Dr. Stan Metzenberg for their support and encouragement and answering every questions. I would like to thank my colleagues in Dr. Oppenheimer’s lab for helping me throughout the project. I would like to express gratitude towards my family for their never ending support and especially would like to thank my girlfriend and my best friend, Shadi Asadabadi for extensive support and patience she has showed in my journey throughout the past two years -

Proceedings of SDAS 1997
Proceedings of the South Dakota Academy of Science Volume 76 1997 Published by the South Dakota Academy of Science Academy Founded November 22, 1915 Editor Kenneth F. Higgins Terri Symens, Wildlife & Fisheries, SDSU provided secretarial assistance Tom Holmlund, Graphic Designer We thank former editor Emil Knapp for compiling the articles contained in this volume. TABLE OF CONTENTS Minutes of the Eighty-Second Annual Meeting of the South Dakota Academy of Science........................................................................................1 Presidential Address: Can we live with our paradigms? Sharon A. Clay ..........5 Complete Senior Research Papers presented at The 82nd Annual Meeting of the South Dakota Academy of Science Fishes of the Mainstem Cheyenne River in South Dakota. Douglas R. Hampton and Charles R. Berry, Jr. ...........................................11 Impacts of the John Morrell Meat Packing Plant on Macroinvertebrates in the Big Sioux River in Sioux Falls, South Dakota. Craig N. Spencer, Gwen Warkenthien, Steven F. Lehtinen, Elizabeth A. Ring, and Cullen R. Robbins ...................................................27 Winter Survival and Overwintering Behavior in South Dakota Oniscidea (Crustacea, Isopoda). Jonathan C. Wright ................................45 Fluctuations in Daily Activity of Muskrates in Eastern South Dakota. Joel F. Lyons, Craig D. Kost, and Jonathan A. Jenks..................................57 Occurrence of Small, Nongame Mammals in South Dakota’s Eastern Border Counties, 1994-1995. Kenneth F. Higgins, Rex R. Johnson, Mark R. Dorhout, and William A. Meeks ....................................................65 Use of a Mail Survey to Present Mammal Distributions in South Dakota. Carmen A. Blumberg, Jonathan A. Jenks, and Kenneth F. Higgins ................................................................................75 A Survey of Natural Resource Professionals Participating in Waterfowl Hunting in South Dakota. Jeffrey S. Gleason and Jonathan A. -

California State University, Northridge the Effects Of
CALIFORNIA STATE UNIVERSITY, NORTHRIDGE THE EFFECTS OF SUGAR ALCOHOLS ON SEA URCHIN GASTRULATION IN LOW CALCIUM SEA WATER A Thesis submitted in partial fulfillment of the requirements For the degree of Master of Science In Biology By Edward Holmes May 2015 Copyright 2015, Edward Holmes ii The thesis of Edward Holmes is approved: _____________________________________ ______________________ Lisa Banner, Ph.D. Date __________________________________ ____________________ Stan Metzenberg, Ph.D. Date __________________________________ ____________________ Steven B. Oppenheimer, Ph.D., Chair Date California State University, Northridge iii DEDICATION This research and thesis project has been dedicated to the Holy Trinity. To my heavenly Abba Father who has adopted me as His son To my Lord and Savior Jesus Christ To the Holy Spirit who is my Comforter and Counselor For their perfect love, grace and mercy For their eternal honor and glory iv ACKNOWLEDGEMENTS Thank you: To Dr. Steven Oppenheimer as a mentor and adviser for your patience, encouragement, guidance, and understanding throughout my research and thesis project. To Dr. Stan Metzenberg for your time and constructive criticism as a thesis committee member. To Dr. Lisa Banner for your time and constructive criticism as a thesis committee member. To my parents Roger and Phyllis and my brothers Jeff and Dwayne for your love, support, generousity, and patience which enabled me to complete my research and thesis project. To my brothers and sisters in Jesus Christ from United Campus Ministry (UCM), the Campus Outreach Response Team (CORT), MT28, and Intervarsity Christian Fellowship for your love, encouragement and prayer support. To my laboratory partners Kathy Fernando and Tiffany Smith for their teamwork. -

Interspecific Variation in Metamorphic Competence in Marine Invertebrates: the Significance for Comparative Investigations Into the Timing of Metamorphosis Cory D
662 Interspecific variation in metamorphic competence in marine invertebrates: the significance for comparative investigations into the timing of metamorphosis Cory D. Bishop,1,* Megan J. Huggett,* Andreas Heyland,y,§ Jason Hodin,¶ and Bruce P. Brandhorst** *Kewalo Marine Laboratories, 41 Ahui St. Honolulu, HI 96813 USA; yFriday Harbor Laboratories University of Washington, 620 University Road, Friday Harbor, WA 98250 USA; §Whitney Laboratory for Marine Biosciences, University of Florida, 9505 Ocean Shore Blvd, FL 32080 USA; ¶Hopkins Marine Station, Stanford University, Oceanview Boulevard, Pacific Grove, CA, 93950 USA; and **Department of Molecular Biology and Biochemistry, Simon Fraser University, Burnaby, BC V5A 1S6, Canada Synopsis Metamorphosis in marine invertebrate larvae is a dynamic, environmentally dependent process that integrates ontogeny with habitat selection. The capacity of many marine invertebrate larvae to survive and maintain metamorphic competence in the absence of environmental cues has been hypothesized to be an adaptive convergence (Hadfield and others 2001). A survey of the literature reveals that a single generalized hypothesis about metamorphic competence as an adaptive convergence is not sufficient to account for interspecific variation in this character. In an attempt to capture this variation, we discuss the “desperate larva hypothesis” and propose two additional hypotheses called the “variable retention hypothesis” and the “death before dishonor hypothesis.” To validate these additional hypotheses we collected data on taxa from the published literature and performed a contingency analysis to detect correlations between spontaneous metamorphosis, habitat specificity and/or larval life-history mode, three characters relevant to environmentally induced settlement and metamorphosis. In order to account for phylogenetic bias in these correlations, we also constructed a phylogeny of these taxa and again performed a character-correlation analysis. -

Sea Urchin Aquaculture
American Fisheries Society Symposium 46:179–208, 2005 © 2005 by the American Fisheries Society Sea Urchin Aquaculture SUSAN C. MCBRIDE1 University of California Sea Grant Extension Program, 2 Commercial Street, Suite 4, Eureka, California 95501, USA Introduction and History South America. The correct color, texture, size, and taste are factors essential for successful sea The demand for fish and other aquatic prod- urchin aquaculture. There are many reasons to ucts has increased worldwide. In many cases, develop sea urchin aquaculture. Primary natural fisheries are overexploited and unable among these is broadening the base of aquac- to satisfy the expanding market. Considerable ulture, supplying new products to growing efforts to develop marine aquaculture, particu- markets, and providing employment opportu- larly for high value products, are encouraged nities. Development of sea urchin aquaculture and supported by many countries. Sea urchins, has been characterized by enhancement of wild found throughout all oceans and latitudes, are populations followed by research on their such a group. After World War II, the value of growth, nutrition, reproduction, and suitable sea urchin products increased in Japan. When culture systems. Japan’s sea urchin supply did not meet domes- Sea urchin aquaculture first began in Ja- tic needs, fisheries developed in North America, pan in 1968 and continues to be an important where sea urchins had previously been eradi- part of an integrated national program to de- cated to protect large kelp beds and lobster fish- velop food resources from the sea (Mottet 1980; eries (Kato and Schroeter 1985; Hart and Takagi 1986; Saito 1992b). Democratic, institu- Sheibling 1988). -

The Biogeography and Community Structure of Kelp Forest Macroinvertebrates Laurel A
Marine Ecology. ISSN 0173-9565 ORIGINAL ARTICLE The biogeography and community structure of kelp forest macroinvertebrates Laurel A. Zahn, Jeremy T. Claisse*, Jonathan P. Williams, Chelsea M. Williams & Daniel J. Pondella II Vantuna Research Group, Occidental College, Los Angeles, CA, USA Keywords Abstract Benthic marine organisms; Channel Islands; invertebrates; kelp beds; marine protected Understanding species distributions and their community structure is increas- areas; Southern California Bight. ingly important when taking an ecosystem-based approach to conservation and management. However, knowledge of the distribution and community Correspondence structure of species in mid-range trophic levels (e.g. macroinvertebrates) is Jeremy T. Claisse, Biological Sciences lacking in most marine ecosystems. Our study aimed to examine the spatial Department, California State Polytechnic distribution and community-level biogeographic patterns of common kelp University, Pomona 3801 West Temple forest–rocky reef macroinvertebrates in Southern California and to evaluate Avenue Pomona, CA 91768, USA. E-mail: [email protected] the effects of environmental gradients on these communities. Quantitative SCUBA surveys were used to estimate macroinvertebrate densities at 92 sites *Present address: Biological Sciences from 2008–2012. Non-metric multidimensional scaling was used to evaluate Department, California State Polytechnic spatial patterns of macroinvertebrate communities among Regions. We found University, Pomona, CA, USA that kelp forest–rocky reef macroinvertebrate communities are distinct among different island and mainland regions, and their community patterns exhib- Accepted: 17 October 2015 ited a strong relationship with an environmental gradient (i.e. sea surface doi: 10.1111/maec.12346 temperature) even after controlling for geographic distance between sites. High abundances of urchin species (Strongylocentrotus purpuratus and Strongylocentrotus franciscanus) were strong drivers of regional differences. -

Marine Ecology Progress Series 457:85
This authors' personal copy may not be publicly or systematically copied or distributed, or posted on the Open Web, except with written permission of the copyright holder(s). It may be distributed to interested individuals on request. Vol. 457: 85–99, 2012 MARINE ECOLOGY PROGRESS SERIES Published June 21 doi: 10.3354/meps09693 Mar Ecol Prog Ser Geographic variation in demography of a temperate reef snail: importance of multiple life-history traits Rebecca G. Martone1,2,*, Fiorenza Micheli1 1Hopkins Marine Station, Stanford University, 120 Oceanview Blvd., Pacific Grove, California 93950, USA 2Present address: Institute for Resources, Environment and Sustainability, The University of British Columbia, Aquatic Ecosystem Research Laboratory, 429-2202 Main Mall, Vancouver, British Columbia V6T 1Z4, Canada ABSTRACT: Individual- and population-level performance may reflect trade-offs between energy allocation to different key demographic processes, such as growth and reproduction, which can, in turn, be influenced by local biotic and abiotic conditions. We explored geographic variation in demographic rates of an exploited benthic species, the wavy-turban snail Megastraea undosa, along the Pacific coast of Baja California, Mexico. We compared key life-history traits (i.e. fecun- dity, size at maturity, growth, and survivorship) of populations existing between 20 and 170 km apart under different conditions of ocean temperature and food availability. Trade-offs between growth and reproduction were evident across this environmental gradient, with higher growth rates in warmer locations leading to lower size-specific investment in gonad production. Because later onset of reproduction in populations from warmer areas was compensated by greater fecun- dity at larger sizes, geographic variation in life-history strategies resulted in similar age-specific reproductive output among different populations. -
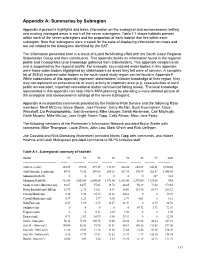
Appendix A: Summaries by Subregion
Appendix A: Summaries by Subregion Appendix A presents highlights and basic information on the ecological and socioeconomic setting and existing managed areas in each of the seven subregions. Table I-1 shows habitats present within each of the seven subregions and the proportion of each habitat that lies within each subregion. Note that subregions were created for the ease of displaying information on maps and are not related to the bioregions identified by the SAT. The information presented here is a result of a joint fact-finding effort with the South Coast Regional Stakeholder Group and their constituents. This appendix builds on information found in the regional profile and incorporates local knowledge gathered from stakeholders. This appendix complements and is supported by the regional profile. For example, key impaired water bodies in this appendix were those water bodies highlighted by stakeholders as areas they felt were of concern. A complete list of 303(d) impaired water bodies in the south coast study region can be found in Appendix F. While subsections of this appendix represent stakeholders’ intimate knowledge of their region, they may not represent an exhaustive list of every activity or important area (e.g. uses/activities at each public access point, important recreational and/or commercial fishing areas). The local knowledge represented in this appendix can help inform MPA planning by providing a more detailed picture of the ecological and socioeconomic settings of the seven subregions. Appendix A incorporates comments provided by the National Park Service and the following RSG members: Merit McCrea, Bruce Steele, Jack Peveler, Gerry Richter, Buck Everingham, Dave Weeshoff, Lia Protopapadakis, Joel Greenberg, Mike Gauger, Sarah Abramson, Carl Mayhugh, Garth Murphy, Mike McCoy, Jack Engle, Norris Tapp, Calla Allison, Mary Jane Foley. -

Influence of Cell Geometry on Division-Plane Positioning
Theory Influence of Cell Geometry on Division-Plane Positioning Nicolas Minc,1,3,4,* David Burgess,2,3 and Fred Chang1,3 1Department of Microbiology and Immunology, Columbia University College of Physicians and Surgeons, 701 W168th Street, New York, NY 10032, USA 2Department of Biology, Boston College, 528 Higgins Hall, 140 Commonwealth Avenue, Chestnut Hill, MA 02167-3811, USA 3Marine Biological Laboratory, 7 MBL Street, Woods Hole, MA 02543, USA 4Present address: Institut Curie, UMR 144 CNRS/IC, 26 rue d’Ulm, 75248 Paris Cedex 05, France *Correspondence: [email protected] DOI 10.1016/j.cell.2011.01.016 SUMMARY from the microtubule (MT) and/or actin cytoskeletons (Grill and Hyman, 2005; Kunda and Baum, 2009; Reinsch and Gonczy, The spatial organization of cells depends on their 1998; Wuhr et al., 2009). Depending on cell type, the division ability to sense their own shape and size. Here, we plane can be set by the orientation of the nucleus during investigate how cell shape affects the positioning of interphase or early prophase or may be modified by rotation or the nucleus, spindle and subsequent cell division movement of the spindle during anaphase. How these force- plane. To manipulate geometrical parameters in generating systems globally sense the shape and dimensions a systematic manner, we place individual sea urchin of the cell remains an outstanding question. The single-cell sea urchin zygote is an attractive cell type for eggs into microfabricated chambers of defined studying the effects of cell geometry. To date, many well-charac- geometry (e.g., triangles, rectangles, and ellipses). -

Echinodermata: the Complex Immune System in Echinoderms
Echinodermata: The Complex Immune System in Echinoderms L. Courtney Smith, Vincenzo Arizza, Megan A. Barela Hudgell, Gianpaolo Barone, Andrea G. Bodnar, Katherine M. Buckley, Vincenzo Cunsolo, Nolwenn M. Dheilly, Nicola Franchi, Sebastian D. Fugmann, Ryohei Furukawa, Jose Garcia-Arraras, John H. Henson, Taku Hibino, Zoe H. Irons, Chun Li, Cheng Man Lun, Audrey J. Majeske, Matan Oren, Patrizia Pagliara, Annalisa Pinsino, David A. Raftos, Jonathan P. Rast, Bakary Samasa, Domenico Schillaci, Catherine S. Schrankel, Loredana Stabili, Klara Stensväg, and Elisse Sutton Echinoderm Life History and Phylogeny Echinoderms are benthic marine invertebrates living in communities ranging from shallow nearshore waters to the abyssal depths. Often members of this phylum are top predators or herbivores that shape and/or control the ecological characteristics All co-authors contributed equally to this chapter and are listed in alphabetical order. L. C. Smith (*) · M. A. Barela Hudgell · K. M. Buckley Department of Biological Sciences, George Washington University, Washington, DC, USA e-mail: [email protected] V. Arizza · G. Barone · D. Schillaci Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, Palermo, Italy A. G. Bodnar Bermuda Institute of Ocean Sciences, St. George’s Island, Bermuda Gloucester Marine Genomics Institute, Gloucester, MA, USA V. Cunsolo Department of Chemical Sciences, University of Catania, Catania, Italy N. M. Dheilly School of Marine and Atmospheric Sciences, Stony Brook University, Stony Brook, NY, USA N. Franchi Department of Biology, University of Padova, Padua, Italy © Springer International Publishing AG, part of Springer Nature 2018 409 E. L. Cooper (ed.), Advances in Comparative Immunology, https://doi.org/10.1007/978-3-319-76768-0_13 410 L. -

Interpreting the Paleozoogeography and Sea Level History of Thermally
Monographs of the Western North American Naturalist Volume 7 8th California Islands Symposium Article 6 9-9-2014 Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California Daniel R. Muhs U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Lindsey T. Groves Natural History Museum of Los Angeles County, [email protected] R. Randall Schumann U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Follow this and additional works at: https://scholarsarchive.byu.edu/mwnan Recommended Citation Muhs, Daniel R.; Groves, Lindsey T.; and Schumann, R. Randall (2014) "Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California," Monographs of the Western North American Naturalist: Vol. 7 , Article 6. Available at: https://scholarsarchive.byu.edu/mwnan/vol7/iss1/6 This Monograph is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Monographs of the Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Monographs of the Western North American Naturalist 7, © 2014, pp. 82–108 INTERPRETING THE PALEOZOOGEOGRAPHY AND SEA LEVEL HISTORY OF THERMALLY ANOMALOUS MARINE TERRACE FAUNAS: A CASE STUDY FROM THE LAST INTERGLACIAL COMPLEX OF SAN CLEMENTE ISLAND, CALIFORNIA Daniel R. Muhs1,3, Lindsey T. Groves2, and R. Randall Schumann1 ABSTRACT.—Marine invertebrate faunas with mixtures of extralimital southern and extralimital northern faunal elements, called thermally anomalous faunas, have been recognized for more than a century in the Quaternary marine terrace record of the Pacific Coast of North America.