The Lysosomal Sialic Acid Transporter Sialin Is Required for Normal CNS Myelination
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neuroradiologic Findings in Fucosidosis, a Rare Lysosomal Storage Disease
Neuroradiologic Findings in Fucosidosis, a Rare Lysosomal Storage Disease James M. Provenzale, Daniel P. Barboriak, and Katherine Sims Summary: Fucosidosis is a rare lysosomal storage disorder with vacuolated secondary lysosomes containing some fine the clinical features of mental retardation, cardiomegaly, dysos- fibrillar material and lamellated membrane structures. tosis multiplex, progressive neurologic deterioration, and early A magnetic resonance (MR) examination at the age of death. The neuroradiologic findings in two patients are reported, 10 years showed confluent regions of hyperintense signal and include abnormalities within the globus pallidus (both pa- on T2-weighted images in the periventricular regions, tients) and periventricular white matter (one patient). most prominent surrounding the atria of the lateral ventri- cles. Hyperintense signal was noted in the globus pallidus Index terms: Brain, metabolism; Degenerative disease, Pediatric on T1-weighted images, with corresponding hypointense neuroradiology signal on T2-weighted images (Fig 1). Fucosidosis is a rare metabolic disorder caused by decreased amounts of the enzyme Case 2 a-L-fucosidase, which results in the accumula- A 2-year-old boy was examined for speech delay and tion of fucose-rich storage products within psychomotor retardation. He was born at term after a many organs, including the brain. Patients with normal pregnancy, and appropriate development oc- this disorder usually have delayed intellectual curred during the first year of life. However, by age 2 years, and motor development, and often die within the patient had not developed speech, exhibited autistic the first decade of life. Computed tomographic behavior, and performed motor tasks poorly. Physical ex- (CT) findings have been reported in a few cases amination findings included coarsened facial features, nar- (1, 2). -

The Concise Guide to Pharmacology 2019/20
Edinburgh Research Explorer THE CONCISE GUIDE TO PHARMACOLOGY 2019/20 Citation for published version: Cgtp Collaborators 2019, 'THE CONCISE GUIDE TO PHARMACOLOGY 2019/20: Transporters', British Journal of Pharmacology, vol. 176 Suppl 1, pp. S397-S493. https://doi.org/10.1111/bph.14753 Digital Object Identifier (DOI): 10.1111/bph.14753 Link: Link to publication record in Edinburgh Research Explorer Document Version: Publisher's PDF, also known as Version of record Published In: British Journal of Pharmacology General rights Copyright for the publications made accessible via the Edinburgh Research Explorer is retained by the author(s) and / or other copyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associated with these rights. Take down policy The University of Edinburgh has made every reasonable effort to ensure that Edinburgh Research Explorer content complies with UK legislation. If you believe that the public display of this file breaches copyright please contact [email protected] providing details, and we will remove access to the work immediately and investigate your claim. Download date: 28. Sep. 2021 S.P.H. Alexander et al. The Concise Guide to PHARMACOLOGY 2019/20: Transporters. British Journal of Pharmacology (2019) 176, S397–S493 THE CONCISE GUIDE TO PHARMACOLOGY 2019/20: Transporters Stephen PH Alexander1 , Eamonn Kelly2, Alistair Mathie3 ,JohnAPeters4 , Emma L Veale3 , Jane F Armstrong5 , Elena Faccenda5 ,SimonDHarding5 ,AdamJPawson5 , Joanna L -

Towards the Elucidation of Orphan Lysosomal Transporters Quentin Verdon
Towards the Elucidation of Orphan Lysosomal Transporters Quentin Verdon To cite this version: Quentin Verdon. Towards the Elucidation of Orphan Lysosomal Transporters. Cancer. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS144. tel-01827233 HAL Id: tel-01827233 https://tel.archives-ouvertes.fr/tel-01827233 Submitted on 2 Jul 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS144 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY PREPAREE A L’UNIVERSITE PARIS-SUD ECOLE DOCTORALE N°568 BIOSIGNE | Signalisations et réseaux intégratifs en biologie Spécialité de doctorat : aspects moléculaires et cellulaires de la biologie Par Mr Quentin Verdon Towards the elucidation of orphan lysosomal transporters: several shots on target and one goal Thèse présentée et soutenue à Paris le 29/06/2016 » : Composition du Jury : Mr Le Maire Marc Professeur, Université Paris-Sud Président Mr Birman Serge Directeur de recherche, CNRS Rapporteur Mr Murray James Assistant professor, Trinity college Dublin Rapporteur Mr Goud Bruno Directeur de recherche, CNRS Examinateur Mr Gasnier Bruno Directeur de recherche, CNRS Directeur de thèse Mme Sagné Corinne Chargée de recherche, INSERM Co-directeur de thèse Table of contents Remerciements (acknowledgements) 6 Abbreviations 7 Abstracts 10 Introduction 12 1 Physiology of lysosomes 12 1.1 Discovery and generalities 12 1.2 Degradative function 13 1.3. -

Toxicological Profile for Nitrate and Nitrite
NITRATE AND NITRITE 29 3. HEALTH EFFECTS 3.1 INTRODUCTION The primary purpose of this chapter is to provide public health officials, physicians, toxicologists, and other interested individuals and groups with an overall perspective on the toxicology of nitrate and nitrite. It contains descriptions and evaluations of toxicological studies and epidemiological investigations and provides conclusions, where possible, on the relevance of toxicity and toxicokinetic data to public health. A glossary and list of acronyms, abbreviations, and symbols can be found at the end of this profile. 3.2 DISCUSSION OF HEALTH EFFECTS BY ROUTE OF EXPOSURE To help public health professionals and others address the needs of persons living or working near hazardous waste sites, the information in this section is organized first by route of exposure (inhalation, oral, and dermal) and then by health effect (e.g., death, systemic, immunological, neurological, reproductive, developmental, and carcinogenic effects). These data are discussed in terms of three exposure periods: acute (14 days or less), intermediate (15–364 days), and chronic (365 days or more). Levels of significant exposure for each route and duration are presented in tables and illustrated in figures. The points in the figures showing no-observed-adverse-effect levels (NOAELs) or lowest- observed-adverse-effect levels (LOAELs) reflect the actual doses (levels of exposure) used in the studies. LOAELs have been classified into "less serious" or "serious" effects. "Serious" effects are those that evoke failure in a biological system and can lead to morbidity or mortality (e.g., acute respiratory distress or death). "Less serious" effects are those that are not expected to cause significant dysfunction or death, or those whose significance to the organism is not entirely clear. -

Sialic Acid Storage Disease
Sialic acid storage disease Description Sialic acid storage disease is an inherited disorder that primarily affects the nervous system. People with sialic acid storage disease have signs and symptoms that may vary widely in severity. This disorder is generally classified into one of three forms: infantile free sialic acid storage disease, Salla disease, and intermediate severe Salla disease. Infantile free sialic acid storage disease (ISSD) is the most severe form of this disorder. Babies with this condition have severe developmental delay, weak muscle tone ( hypotonia), and failure to gain weight and grow at the expected rate (failure to thrive). They may have unusual facial features that are often described as "coarse," seizures, bone malformations, an enlarged liver and spleen (hepatosplenomegaly), and an enlarged heart (cardiomegaly). The abdomen may be swollen due to the enlarged organs and an abnormal buildup of fluid in the abdominal cavity (ascites). Affected infants may have a condition called hydrops fetalis in which excess fluid accumulates in the body before birth. Children with this severe form of the condition usually live only into early childhood. Salla disease is a less severe form of sialic acid storage disease. Babies with Salla disease usually begin exhibiting hypotonia during the first year of life and go on to experience progressive neurological problems. Signs and symptoms of Salla disease include intellectual disability and developmental delay, seizures, problems with movement and balance (ataxia), abnormal tensing of the muscles (spasticity), and involuntary slow, sinuous movements of the limbs (athetosis). Individuals with Salla disease usually survive into adulthood. People with intermediate severe Salla disease have signs and symptoms that fall between those of ISSD and Salla disease in severity. -

Sialin (SLC17A5) Functions As a Nitrate Transporter in the Plasma Membrane
Sialin (SLC17A5) functions as a nitrate transporter in the plasma membrane Lizheng Qina,1, Xibao Liub,1, Qifei Suna, Zhipeng Fana, Dengsheng Xiaa, Gang Dinga, Hwei Ling Ongb, David Adamsc, William A. Gahlc, Changyu Zhengb, Senrong Qia, Luyuan Jina, Chunmei Zhanga, Liankun Gud, Junqi Hee, Dajun Dengd,2, Indu S. Ambudkarb,2, and Songlin Wanga,e,2 aSalivary Gland Disease Center and Beijing Key Laboratory of Tooth Regeneration and Function Reconstruction, Capital Medical University School of Stomatology, Beijing 100050, China; bMolecular Physiology and Therapeutics Branch, National Institute of Dental and Craniofacial Research, Bethesda, MD 20892; cMedical Genetics Branch, National Human Genome Research Institute, Bethesda, MD 20892; dKey Laboratory of Carcinogenesis and Translational Research, Peking University Cancer Hospital and Institute, Beijing 100142, China; and eDepartment of Biochemistry and Molecular Biology, Capital Medical University School of Basic Medicine, Beijing 100069, China Edited by Mark Gladwin, University of Pittsburg Medical Center, Pittsburgh, PA and accepted by the Editorial Board June 5, 2012 (received for review October 13, 2011) In vivo recycling of nitrate (NO −) and nitrite (NO −) is an important Nitrate uptake into salivary glands represents the key initial step 3 2 − alternative pathway for the generation of nitric oxide (NO) and in NO clearance from the serum; however, the mechanism me- 3 − maintenance of systemic nitrate–nitrite–NO balance. More than diating transport of NO3 in salivary gland epithelial cells has not − − fl 25% of the circulating NO3 is actively removed and secreted by yet been established. In this study, we examined NO3 in ux in − + salivary glands. Oral commensal bacteria convert salivary NO3 to salivary gland cells. -

Program Download
5TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE ROME, ITALY NOVEMBER 1-4 2017 EMBRACING INNOVATION ADVANCING THE CURE ISMRD would like to say a very special thank you to the following organizations and companies who have very generously given donations to support the 5th International Conference on Glycoproteinoses. THE WAGNER FOUNDATION ISMRD is very grateful for all the help and support that Symposia has given us in the organization of our Conference on-the-ground support in Rome. Dedicated to helping patients in the rare disease community with unmet medical needs Ultragenyx Pharmaceutical Inc. is a clinical-stage biopharmaceutical company committed to creating new therapeutics to combat serious, debilitating diseases. www.ultragenyx.com © 2017 Ultragenyx Pharmaceutical Inc. www.ultragenyx.com 8/16 MRCP-UGNX-00008 5TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE 2017 Welcome to Rome! On behalf of ISMRD, I would like to welcome Of course, our conference would not take you all to the Fifth Glycoproteinoses place without significant charitable donations. International Conference on 'Embracing I would like to thank the following companies Innovation and Advancing the Cure'. and foundations, Ultragenyx, EveryLife Foundation, Sanofi/Genzyme, Amicus and The ISMRD is thrilled to bring our International Wagner Foundation. I would also like to extend Conference to Europe allowing us to connect our very grateful thanks to Symposia who are with families, researchers, clinicians, support event planners, who have worked alongside us groups and others who work in the field of rare here in Rome and have helped you check in at diseases. But, more importantly to help make registration. They will be on site throughout the the invaluable connections for families who meeting offering support and answering any perhaps have never met another family with questions you may have. -

Potential Roles of Nitrate and Nitrite in Nitric Oxide Metabolism in the Eye Ji Won Park1, Barbora Piknova1, Audrey Jenkins2, David Hellinga2, Leonard M
www.nature.com/scientificreports OPEN Potential roles of nitrate and nitrite in nitric oxide metabolism in the eye Ji Won Park1, Barbora Piknova1, Audrey Jenkins2, David Hellinga2, Leonard M. Parver3 & Alan N. Schechter1* Nitric oxide (NO) signaling has been studied in the eye, including in the pathophysiology of some eye diseases. While NO production by nitric oxide synthase (NOS) enzymes in the eye has been − characterized, the more recently described pathways of NO generation by nitrate ( NO3 ) and nitrite − (NO2 ) ions reduction has received much less attention. To elucidate the potential roles of these pathways, we analyzed nitrate and nitrite levels in components of the eye and lacrimal glands, primarily in porcine samples. Nitrate and nitrite levels were higher in cornea than in other eye parts, while lens contained the least amounts. Lacrimal glands exhibited much higher levels of both ions compared to other organs, such as liver and skeletal muscle, and even to salivary glands which are known to concentrate these ions. Western blotting showed expression of sialin, a known nitrate transporter, in the lacrimal glands and other eye components, and also xanthine oxidoreductase, a nitrate and nitrite reductase, in cornea and sclera. Cornea and sclera homogenates possessed a measurable amount of nitrate reduction activity. These results suggest that nitrate ions are concentrated in the lacrimal glands by sialin and can be secreted into eye components via tears and then reduced to nitrite and NO, thereby being an important source of NO in the eye. Te NO generation pathway from L-arginine by endogenous NOS enzymes under normoxic conditions has been central in identifying the physiological roles of NO in numerous biological processes1. -

Fast Urinary Screening of Oligosaccharidoses by MALDI-TOF/TOF Mass Spectrometry
Fast urinary screening of oligosaccharidoses by MALDI-TOF/TOF mass spectrometry. Laurent Bonesso, Monique Piraud, Céline Caruba, Emmanuel van Obberghen, Raymond Mengual, Charlotte Hinault To cite this version: Laurent Bonesso, Monique Piraud, Céline Caruba, Emmanuel van Obberghen, Raymond Mengual, et al.. Fast urinary screening of oligosaccharidoses by MALDI-TOF/TOF mass spectrometry.. Orphanet Journal of Rare Diseases, BioMed Central, 2014, 9 (1), pp.19. 10.1186/1750-1172-9-19. inserm- 00945684 HAL Id: inserm-00945684 https://www.hal.inserm.fr/inserm-00945684 Submitted on 12 Feb 2014 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Bonesso et al. Orphanet Journal of Rare Diseases 2014, 9:19 http://www.ojrd.com/content/9/1/19 RESEARCH Open Access Fast urinary screening of oligosaccharidoses by MALDI-TOF/TOF mass spectrometry Laurent Bonesso1, Monique Piraud5, Céline Caruba1, Emmanuel Van Obberghen1,2,3,4, Raymond Mengual1† and Charlotte Hinault1,2,3,4*† Abstract Background: Oligosaccharidoses, which belong to the lysosomal storage diseases, are inherited metabolic disorders due to the absence or the loss of function of one of the enzymes involved in the catabolic pathway of glycoproteins and indirectly of glycosphingolipids. -

4Th Glycoproteinoses International Conference Advances in Pathogenesis and Therapy
Program & Abstracts 4TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE ADVANCES IN PATHOGENESIS AND THERAPY ISMRD ST. LOUIS, MISSOURI, UNITED STATES Program & Abstracts I SM R D ADVANCES IN PATHOGENESIS AND THERAPY Program & Abstracts ISMRD would like to say A Very Special Thank You to the following organizations and companies who have very generously given donations and sponsorship to support the 4th International Conference on Glycoproteinoses THE PRENILLE EDWARD MALLINCKRODT FOUNDATION JR FOUNDATION MARK HASKINS I SM R D 4TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE 2015 ADVANCES IN PATHOGENESIS AND THERAPY Program & Abstracts ISMRD is very proud to display 10 featured Expression of Hope artworks to be Auctioned at the Gala Dinner. These beautiful prints are from Genzyme’s featured Artwork selection. Contents Welcome 1 SCIENTIFIC COMMITTEE: Stuart Kornfeld ISMRD Mission & Governance 3 (Chair, Scientifi c Planning Committee) Steve Walkley Sara Cathey ISMRD General Information 5 Richard Steet Sean Thomas Ackley, Philippines Miriam Storchli, Switzerland Alessandra d’Azzo ‘Hope’ by Sarah Noble, New Zealand Scientifi c Program 9 FAMILY CONFERENCE COMMITTEE: Family Program for Mucolipidosis 11 Jenny Noble (Conference Organiser) Jackie James (Conference Organiser Family Program For Alpha Mannosidosis /Sialidosis/ 13 - St. Louis) Fucosidosis/Aspartylglucosaminuria Mark Stark John Forman ‘All around the world’ by Zih Yun Li , Taiwan Childrens Program 16 Susan Kester Carolyn Paisley-Dew Tish Adkins Abstracts 17 Sara DeAngelis, Russia Gayle Rose, United States Speaker Profi les 60 Delegates 81 Helen Walker, Australia Nicklas Harkins, Canada Naomi Arai, Japan David Wentworth, Serbia I SM R D 4TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE 2015 ADVANCES IN PATHOGENESIS AND THERAPY Program & Abstracts On behalf of the Scientifi c Planning Committee, I want to extend a warm welcome to all the investigators and Welcome! families who have traveled to St. -

International Conference
5TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE Rome, Italy November 1-4 2017 EMBRACING INNOVATION ADVANCING THE CURE PROGRAM & ABSTRACTS 5TH GLYCOPROTEINOSES INTERNATIONAL CONFERENCE ROME, ITALY NOVEMBER 1-4 2017 EMBRACING INNOVATION ADVANCING THE CURE ISMRD would like to say a very special thank you to the following organizations and companies who have very generously given donations to support the 5th International Conference on Glycoproteinoses. ISMRD is an internationally focused not-for-profi t organization whose mission is to advocate for families and patients aff ected by one of the following disorders. Alpha-Mannosidosis THE WAGNER FOUNDATION Aspartylglucosaminuria Beta-Mannosidosis Fucosidosis Galactosialidosis ISMRD is very grateful for all the help and support that Symposia has given us Sialidosis (Mucolipidosis I) in the organization of our Conference on-the-ground support in Rome. Mucolipidosis II, II/III, III alpha/beta Mucolipidosis III Gamma Schindler Disease EMBRACING INNOVATION ADVANCING THE CURE SCIENTIFIC COMMITTEE: Alessandra d’Azzo CHAIR Contents Amelia Morrone Italy Richard Steet USA Welcome 2 Heather Flanagan-Steet USA ISMRD Mission & Governance 4 Dag Malm Norway ISMRD General Information 6 Thomas Braulke Dedicated to helping patients Germany in the rare disease community Stuart Kornfeld with unmet medical needs Scientifi c Program 10 USA Ultragenyx Pharmaceutical Inc. is a clinical-stage Family Program 14 ISMRD CONFERENCE biopharmaceutical company committed to creating new COMMITTEE: therapeutics to combat serious, -
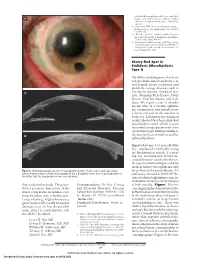
Mucolipidosis Type I)
endothelial keratoplasty in 200 eyes: early chal- A lenges and techniques to enhance donor adherence. J Cataract Refract Surg. 2006;32(3): 411-418. 4. Culbertson WW. Descemet stripping endothe- lial keratoplasy. Int Ophthalmol Clin. 2006;46 (3):155-168. 5. Ellis DR, Cohen KL. Sulfur hexafluoride gas in the repair of Descemet’s membrane detachment. Cornea. 1995;14(4):436-437. 6. Sorenson A. DSEK prolene pull. http://www .newmediamedicine.com/videos/2007/01/17 /dsek-prolene-pull-andrew-sorenson-md/. Ac- cessed March 18, 2007. Cherry Red Spot in Sialidosis (Mucolipidosis Type I) The differential diagnosis of a cherry red spot in the macula includes cen- tral retinal artery occlusion and metabolic storage diseases such as B Tay-Sachs disease, Sandhoff dis- ease, Niemann-Pick disease, Fabry disease, Gaucher disease, and siali- dosis. We report a case of an ado- lescent who, at a routine ophthal- mic examination, was found to have a cherry red spot in the maculae of both eyes. Laboratory investigation results showed that the patient had mucolipidosis type I, which is a rare lysosomal storage disease with clini- cal and histologic findings similar to C the mucopolysaccharidoses and the sphingolipidoses. Report of a Case. A 14-year-old white boy complained of difficulty seeing the blackboard at school. A screen- ing eye examination found de- creased distance vision in both eyes. He was of normal intelligence and his medical history was significant only Figure 3. Slitlamp photograph of case 3 at postoperative month 1 shows a clear cornea (A), and on for scoliosis and seasonal allergies.