Holostei, Parasemionotidae), a Fossil Fish from the Continental Middle Jurassic of Congo (D
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

BONY FISHES 602 Bony Fishes
click for previous page BONY FISHES 602 Bony Fishes GENERAL REMARKS by K.E. Carpenter, Old Dominion University, Virginia, USA ony fishes constitute the bulk, by far, of both the diversity and total landings of marine organisms encoun- Btered in fisheries of the Western Central Atlantic.They are found in all macrofaunal marine and estuarine habitats and exhibit a lavish array of adaptations to these environments. This extreme diversity of form and taxa presents an exceptional challenge for identification. There are 30 orders and 269 families of bony fishes presented in this guide, representing all families known from the area. Each order and family presents a unique suite of taxonomic problems and relevant characters. The purpose of this preliminary section on technical terms and guide to orders and families is to serve as an introduction and initial identification guide to this taxonomic diversity. It should also serve as a general reference for those features most commonly used in identification of bony fishes throughout the remaining volumes. However, I cannot begin to introduce the many facets of fish biology relevant to understanding the diversity of fishes in a few pages. For this, the reader is directed to one of the several general texts on fish biology such as the ones by Bond (1996), Moyle and Cech (1996), and Helfman et al.(1997) listed below. A general introduction to the fisheries of bony fishes in this region is given in the introduction to these volumes. Taxonomic details relevant to a specific family are explained under each of the appropriate family sections. The classification of bony fishes continues to transform as our knowledge of their evolutionary relationships improves. -

From the Crato Formation (Lower Cretaceous)
ORYCTOS.Vol. 3 : 3 - 8. Décembre2000 FIRSTRECORD OT CALAMOPLEU RUS (ACTINOPTERYGII:HALECOMORPHI: AMIIDAE) FROMTHE CRATO FORMATION (LOWER CRETACEOUS) OF NORTH-EAST BRAZTL David M. MARTILL' and Paulo M. BRITO'z 'School of Earth, Environmentaland PhysicalSciences, University of Portsmouth,Portsmouth, POl 3QL UK. 2Departmentode Biologia Animal e Vegetal,Universidade do Estadode Rio de Janeiro, rua SâoFrancisco Xavier 524. Rio de Janeiro.Brazll. Abstract : A partial skeleton representsthe first occurrenceof the amiid (Actinopterygii: Halecomorphi: Amiidae) Calamopleurus from the Nova Olinda Member of the Crato Formation (Aptian) of north east Brazil. The new spe- cimen is further evidencethat the Crato Formation ichthyofauna is similar to that of the slightly younger Romualdo Member of the Santana Formation of the same sedimentary basin. The extended temporal range, ?Aptian to ?Cenomanian,for this genus rules out its usefulnessas a biostratigraphic indicator for the Araripe Basin. Key words: Amiidae, Calamopleurus,Early Cretaceous,Brazil Première mention de Calamopleurus (Actinopterygii: Halecomorphi: Amiidae) dans la Formation Crato (Crétacé inférieur), nord est du Brésil Résumé : la première mention dans le Membre Nova Olinda de la Formation Crato (Aptien ; nord-est du Brésil) de I'amiidé (Actinopterygii: Halecomorphi: Amiidae) Calamopleurus est basée sur la découverted'un squelettepar- tiel. Le nouveau spécimen est un élément supplémentaireindiquant que I'ichtyofaune de la Formation Crato est similaire à celle du Membre Romualdo de la Formation Santana, située dans le même bassin sédimentaire. L'extension temporelle de ce genre (?Aptien à ?Cénomanien)ne permet pas de le considérer comme un indicateur biostratigraphiquepour le bassin de l'Araripe. Mots clés : Amiidae, Calamopleurus, Crétacé inférieu4 Brésil INTRODUCTION Araripina and at Mina Pedra Branca, near Nova Olinda where cf. -

Blackchin Tilapia (Sarotherodon Melanotheron) Ecological Risk Screening Summary
U.S. Fish and Wildlife Service Blackchin Tilapia (Sarotherodon melanotheron) Ecological Risk Screening Summary Web Version – 10/01/2012 Photo: © U.S. Geological Survey From Nico and Neilson (2014). 1 Native Range and Nonindigenous Occurrences Native Range From Nico and Neilson (2014): “Tropical Africa. Brackish estuaries and lagoons from Senegal to Zaire (Trewavas 1983).” Nonindigenous Occurrences From Nico and Neilson (2014): “Established in Florida and Hawaii. Evidence indicates it is spreading rapidly in both fresh and salt water around island of Oahu, Hawaii (Devick 1991b).” “The first documented occurrence of this species in Florida was a specimen gillnetted by commercial fishermen in Hillsborough Bay near Tampa, Hillsborough County, in 1959 (Springer and Finucane 1963). Additional records for the western part of the state indicate that this species is established in brackish and freshwaters in eastern Tampa Bay and in adjoining drainages in Hillsborough County, ranging from the Alafia River south to Cockroach Bay. The species has been recorded from the Alafia River from its mouth up to Lithia Springs; from the Hillsborough River, Bullfrog Creek, the Palm River, and the Little Manatee River; and from various western drainage and irrigation ditches (Springer and Finucane 1963; Finucane and Rinckey 1967; Buntz Sarotherodon melanotheron Ecological Risk Screening Summary U.S. Fish and Wildlife Service – Web Version – 10/01/2012 and Manooch 1969; Lachner et al. 1970; Courtenay et al. 1974; Courtenay and Hensley 1979; Courtenay and Kohler 1986; Lee et al. 1980 et seq.; Courtenay and Stauffer 1990; DNR collections; UF museum specimens). There are two records of this species from the west side of Tampa Bay, in Pinellas County: a collection from Lake Maggiore in St. -

The Early Triassic Jurong Fish Fauna, South China Age, Anatomy, Taphonomy, and Global Correlation
Global and Planetary Change 180 (2019) 33–50 Contents lists available at ScienceDirect Global and Planetary Change journal homepage: www.elsevier.com/locate/gloplacha Research article The Early Triassic Jurong fish fauna, South China: Age, anatomy, T taphonomy, and global correlation ⁎ Xincheng Qiua, Yaling Xua, Zhong-Qiang Chena, , Michael J. Bentonb, Wen Wenc, Yuangeng Huanga, Siqi Wua a State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan), Wuhan 430074, China b School of Earth Sciences, University of Bristol, BS8 1QU, UK c Chengdu Center of China Geological Survey, Chengdu 610081, China ARTICLE INFO ABSTRACT Keywords: As the higher trophic guilds in marine food chains, top predators such as larger fishes and reptiles are important Lower Triassic indicators that a marine ecosystem has recovered following a crisis. Early Triassic marine fishes and reptiles Fish nodule therefore are key proxies in reconstructing the ecosystem recovery process after the end-Permian mass extinc- Redox condition tion. In South China, the Early Triassic Jurong fish fauna is the earliest marine vertebrate assemblage inthe Ecosystem recovery period. It is constrained as mid-late Smithian in age based on both conodont biostratigraphy and carbon Taphonomy isotopic correlations. The Jurong fishes are all preserved in calcareous nodules embedded in black shaleofthe Lower Triassic Lower Qinglong Formation, and the fauna comprises at least three genera of Paraseminotidae and Perleididae. The phosphatic fish bodies often show exceptionally preserved interior structures, including net- work structures of possible organ walls and cartilages. Microanalysis reveals the well-preserved micro-structures (i.e. collagen layers) of teleost scales and fish fins. -

Southampton French Quarter 1382 Specialist Report Download E2: Fish Bone
Southampton French Quarter SOU1382 Specialist Report Download E2 Southampton French Quarter 1382 Specialist Report Download E2: Fish Bone By Rebecca Nicholson Introduction An assemblage of almost 7500 identifiable fish bones was recovered both by hand retrieval during the excavation, but predominantly from the sorted residues of the processed bulk soil samples. During excavations at Southampton French Quarter a total of 188 bulk samples were sieved to 0.5mm (occasionally 1mm) as part of the flotation process for the recovery of plant and animal remains. The sampling strategy followed during the excavation involved, where possible, the full sampling of one rubbish pit and one latrine pit per tenement for each major period, avoiding intercut features or those clearly containing residual material. Occupation surfaces and other distinct features such as hearths were also sampled. Mixed contexts or contexts of uncertain provenance were avoided. While complete standardisation of sample volumes was not possible, wherever practicable samples were 40 litres. Following assessment of the fish remains recovered largely from the > 4mm residues, the richer assemblages were targeted for further fine residue and flot sorting. This report comprises an analysis of all the identified the fish remains from these samples, together with the material collected by hand on site. Methodology The residues from all of the bulk-sieved samples were sorted to 4mm. Where samples were identified as having significant numbers of fish bones, residues were sorted to 2mm. Samples from the Late Saxon deposits which produced fish remains were routinely sorted to 2mm even where fish remains were not abundant, in order to avoid a perceived bias against the recovery of small fish in pre-medieval deposits (see Barrett et al. -
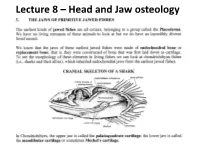
Lecture 8 – Head and Jaw Osteology
Lecture 8 – Head and Jaw osteology More derived fishes (Ray finned fishes) The variability of the jaw structure of bony fishes provides an explanation for the extensive adaptive radiation in the group and why they are so diverse and occupy almost every aquatic niche available. Skull diversity (A) carp, Cyprinus carpio, (B) vampire characin, Hydrolycus scomberoides, (C) catfish Arius felis. (D) cod Gadus morhua. (E) large-mouth bass, Micropterus salmoides (F) The parrotfish Scarus guacamaia. Scale bar = 10 mm WESTNEAT 2004 From an evolutionary standpoint, fishes were the first animals to develop bony jaws. Versatile jaws and multiple feeding strategies allowed fishes to fill, or radiate into, a diverse range of niches. They have evolved to feed in all possible ways – sucking, biting, scraping, nipping, crushing etc. The head of a teleost has 5 main regions: Cranium, jaws, cheeks, hydroid arch, opercula. The head of a fish has five main regions • 1) The CRANIUM is composed of the bones providing direct support and protection to the brain and the visual, Anterior Posterior olfactory, and auditory organs. Below the cranium is the parashenoid bone. • Parasphenoid plays a role in the jaws as Features of the neurocranium sensu lato (from Caranx it acts as a hard melampygus, lateral aspect, left, and posterior aspect, right). A = prevomer, B = ethmoid, C = frontal, D = palate supraoccipital, E = pterotic, F = exoccipital, G = basioccipital, H = foramen magnum, I = parasphenoid, J = orbit. The five main regions Bowfin 2) The JAWS • Lower Jaw – has an Angular articular and dentary bone • Angular articular- The paired bones form the posterior part of either side of the lower jaw and articulate with the suspensorium. -

Marine Early Triassic Osteichthyes from Spiti, Indian Himalayas
Swiss J Palaeontol (2016) 135:275–294 DOI 10.1007/s13358-015-0098-6 Marine Early Triassic Osteichthyes from Spiti, Indian Himalayas 1 1 1 1 Carlo Romano • David Ware • Thomas Bru¨hwiler • Hugo Bucher • Winand Brinkmann1 Received: 12 March 2015 / Accepted: 11 August 2015 / Published online: 28 September 2015 Ó Akademie der Naturwissenschaften Schweiz (SCNAT) 2015 Abstract A new, marine osteichthyan (bony fish) fauna strata of other localities. The study of Early Triassic fish from the Early Triassic of northern India is presented. The assemblages, including the presented one, is fundamental material was collected in situ at localities within Pin Valley for our understanding of the great osteichthyan diversifi- (Lahaul and Spiti District, Himachal Pradesh, India) and is cation after the Late Permian mass extinction event. dated as middle-late Dienerian (one specimen possibly earliest Smithian). The new ichthyofauna includes a lower Keywords Neotethys Á Northern Indian Margin Á jaw of the predatory basal ray-finned fish Saurichthys,a Gondwana Á Anoxia Á Biotic recovery Á Urohyal nearly complete specimen of a parasemionotid neoptery- gian (cf. Watsonulus cf. eugnathoides), as well as further Abbreviations articulated and disarticulated remains (Actinopterygii CMNFV Canadian Museum of Nature (Fossil indet., Actinistia indet.), and thus comprises the most Vertebrate), Ottawa, Canada complete Triassic fish fossils known from the Indian sub- MNHN.F Muse´um National d’Histoire Naturelle, Paris, continent. Saurichthys is known from many Triassic France localities and reached a global distribution rapidly after the PIMUZ Pala¨ontologisches Institut und Museum, Late Permian mass extinction event. Parasemionotidae, a Universita¨tZu¨rich, Zu¨rich, Schweiz species-rich family restricted to the Early Triassic, also achieved widespread distribution during this epoch. -

Marine Fish Osteology a Manual for Archaeologists
Marine Fish Osteology A Manual for Archaeologists f t n Debbi Yee Cannon Department of Archaeology Simon Fraser University Publication no. 18 Burnaby, B.C. 1987 Archaeology Press Simon Fraser University Burnaby, B.C. PUBLICATIONS COMMITTEE Roy L. Carlson (Chairman) Knut R. Fladmark Brian Hayden Philip M. Hobler Jack D. Nance Erie Nelson All rights reserved. No part o f this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording or any information storage and retrieval system, without permission in writing from the publisher. ISBN 0 - 86491-083-5 PRINTED IN CANADA The Department o f Archaeology publishes papers and monographs which relate to its teaching and research interests. Communications concerning publications should be directed to the Chairman o f the Publications Committee. © Copyright 1987 Department of Archaeology Simon Fraser University Marine Fish Osteology A Manual for Archaeologists Debbi Yee Cannon iii Table of Contents A cknow ledgem ents.............................................................................................................. v Introduction........................................................................................................................... 1 Fish Identification................................................................................................................ 1 Fish Rem ains in A rch aeology.......................................................................................... 3 Scope o -

A Redescription of the Lungfish Eoctenodus Hills 1929, with Reassessment of Other Australian Records of the Genus Dipterns Sedgwick & Murchison 1828
Ree. West. Aust. Mus. 1987, 13 (2): 297-314 A redescription of the lungfish Eoctenodus Hills 1929, with reassessment of other Australian records of the genus Dipterns Sedgwick & Murchison 1828. J.A. Long* Abstract Eoctenodus microsoma Hills 1929 (= Dipterus microsoma Hills, 1931) from the Frasnian Blue Range Formation, near Taggerty, Victoria, is found to be a valid genus, differing from Dipterus, and other dipnoans, by the shape of the parasphenoid and toothplates. The upper jaw toothp1ates and entopterygoids, parasphenoid, c1eithrum, anoc1eithrum and scales of Eoctenodus are described. Eoctenodus may represent the earliest member of the Ctenodontidae. Dipterus cf. D. digitatus. from the Late Devonian Gneudna Formation, Western Australia (Seddon, 1969), is assigned to Chirodipterus australis Miles 1977; and Dipterus sp. from the Late Devonian of Gingham Gap, New South Wales (Hills, 1936) is thought to be con generic with a dipnoan of similar age from the Hunter Siltstone, New South Wales. This form differs from Dipterus in the shape of the parasphenoid. The genus Dipterus appears to be restricted to the Middle-Upper Devonian of Europe, North America and the USSR (Laurasia). Introduction Although Hills (1929) recognised a new dipnoan, Eoctenodus microsoma, in the Late Devonian fish remains from the Blue Range Formation, near Taggerty, he later (Hills 1931) altered the generic status of this species after a study trip to Britain in which D,M.S. Watson pointed out similarities between the Australian form and the British genus Dipterus Sedgwick and Murchison 1828. Studies of the head of Dipterus by Westoll (1949) and White (1965) showed the structure of the palate and, in particular, the shape of the parasphenoid which differs from that in the Taggerty dipnoan. -

The Genus Furo (Pisces, Halecomorphi) from the Upper Jurassic Plattenkalke of Germany
ORYCTOS,Vol. 1 :23-35,Octobre 1998 THEGENUS FURO (PISCES, HALECOMORPHI) FROMTHE UPPERJURASSIC PLATTENKALKE OF GERMANY Paul H. LAMBERS PaleontologischeWerkkamer, Biologisch Centrum RUG, Postbus 14,9750 AA Haren, the Netherlands. e-mail: phlambers@ biol.rug.nl Abstract : An overview of the speciesassigned to the genus Furo fowd in the German lithographic limestones of the Solnhofen-area(Bavaria) and Nusplingen (Baden-Wiirttemberg) is presentedand the monophyly of the Upper JurassicFuro is discussed.Six speciescan be recognized: 'F.' latim.anus,'F.' longiserratus, 'F.' microlepidotes, 'E' aldingeri, 'F.' angustus and'F.' miinsteri. Among these 'E' angustus and'F.' miinsteri form a monophyletic group, to which 'F.' aldingeri might be related as well. 'F.' longiserrarus might be closely related to the Ophiopsidae,whereas 'E' microlepidotes shows similarities with the Caturidae. The position of 'F.' latimanus remains to be determined. There are no indications of a monophyletic genusof Furo and the relationshipsof the Upper Jurassicfurids with the Lower Jurassicspecies of Furo remain to be examined. Key words: Eugnathus, Furo, Halecomorphi, phylogeny, Plattenknlke, Tithonian Le genreFuro (Pisces,Halecomorphi) du Jurassiquesupérieur d'Allemagne. Résumé : Les différentes espècesdu genre Furo enprovenancedes gisementsallemands à calcaireslithographiques des régions de Solnhofen (Bavière) et de Nusplingen (Bade-Wiirttemberg)sont présentéeset la monophylie du genre Furo du Jurassiquesupérieur est discutée.Six espècespeuvent être reconnues: '.8' Iatimanus, -

Halecomorphi, Amiidae
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Open Marine Archive BULLETIN DE L’INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 80: 163-170, 2010 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 80: 163-170, 2010 First fossil record of an amiid fish (Halecomorphi, Amiidae) from the Latest Cretaceous of Patagonia, Argentina, and comments on the status of Pappichthys patagonica AMEGHINO , 1906 (Teleostei, Osteoglossidae) by Sergio BOGAN, Louis TAVERNE & Federico L. AGNOLIN BOG A N , S., TAVERNE , L. & AGNO L IN , F.L., 2010 – First fossil Introduction record of an amiid fish (Halecomorphi, Amiidae) from the Latest Cretaceous of Patagonia, Argentina, and comments on the The Amiiformes are halecomorph fishes represented status of Pappichthys patagonica AMEGHINO , 1906 (Teleostei, Osteoglossidae). Bulletin de l’Institut royal des Sciences naturelles today by the single extant species Amia calva de Belgique, Sciences de la Terre, 80: 163-170, 4 figs, Brussels, LINN A EUS , 1766, the bowfin, which is geographically October 31, 2010 – ISSN 0374-6291. distributed among freshwater lakes and rivers in Eastern North America (NE L SON , 2006: 99). Abstract This large predaceous fish constitutes a relict of a taxonomic group widely distributed among most We describe the first authenticated fossil record for the family continents during the Mesozoic and the Caenozoic. Amiidae in Argentina. The specimen consists on an isolated dentary coming from the Uppermost Cretaceous Allen Formation, from Río The first record for the Amiiformes occurs in the Late Negro province, Patagonia, Argentina, and belonging probably Triassic (Norian), whereas the oldest record of the to the genus Amia. -

Hrísheimar: Fish Consumption Patterns
City University of New York (CUNY) CUNY Academic Works School of Arts & Sciences Theses Hunter College Fall 1-3-2019 Hrísheimar: Fish Consumption Patterns Wendi K. Coleman CUNY Hunter College How does access to this work benefit ou?y Let us know! More information about this work at: https://academicworks.cuny.edu/hc_sas_etds/376 Discover additional works at: https://academicworks.cuny.edu This work is made publicly available by the City University of New York (CUNY). Contact: [email protected] Hrísheimar: Fish Consumption Patterns By Wendi K. Coleman Submitted in partial fulfillment of the requirements for the degree of Master of Arts Anthropology, Hunter College The City University of New York 2019 Thesis Sponsor: January 3, 2019_______ Thomas McGovern_____________________ Date Signature January 3, 2019_______ George Hambrecht______________________ Date Signature of Second Reader i ACKNOWLEDGEMENTS There are several people that I would like to thank for encouraging and assisting me in the completion of this thesis. First, I would like to thank my thesis advisor Dr. Thomas McGovern for enthusiastically sharing his vast knowledge on Icelandic archaeological sites and for his support throughout the entire process. I would like to thank my second reader Dr. George Hambrecht for his support. I would also like to thank several members of the NABO lab community including Frank Feeley who taught me how to identify Gadidae species and their bones as well as provided several key resources for me to utilize. I would also like to thank Megan Hicks who spent several hours teaching me to identify Salmonidae species. Frank and Megan were always willing to aid with any odd bones or species questions.