Dudley Edwin Pinnock, B. Sc. (Lond.)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lepidoptera, Zygaenidae
©Ges. zur Förderung d. Erforschung von Insektenwanderungen e.V. München, download unter www.zobodat.at _______Atalanta (Dezember 2003) 34(3/4):443-451, Würzburg, ISSN 0171-0079 _______ Natural enemies of burnets (Lepidoptera, Zygaenidae) 2nd Contribution to the knowledge of hymenoptera paraziting burnets (Hymenoptera: Braconidae, Ichneumonidae, Chaleididae) by Tadeusz Kazmierczak & J erzy S. D ^browski received 18.VIII.2003 Abstract: New trophic relationships between Braconidae, Ichneumonidae, Chaleididae, Pteromalidae, Encyrtidae, Torymidae, Eulophidae (Hymenoptera) and burnets (Lepidoptera, Zygaenidae) collected in selected regions of southern Poland are considered. Introduction Over 30 species of insects from the family Zygaenidae (Lepidoptera) occur in Central Europe. The occurrence of sixteen of them was reported in Poland (D/^browski & Krzywicki , 1982; D/^browski, 1998). Most of these species are decidedly xerothermophilous, i.e. they inhabit dry, open and strongly insolated habitats. Among the species discussed in this paperZygaena (Zygaena) angelicae O chsenheimer, Z. (Agrumenia) carniolica (Scopoli) and Z (Zygaena) loti (Denis & Schiffermuller) have the greatest requirements in this respect, and they mainly live in dry, strongly insolated grasslands situated on lime and chalk subsoil. The remaining species occur in fresh and moist habitats, e. g. in forest meadows and peatbogs. Due to overgrowing of the habitats of these insects with shrubs and trees as a result of natural succession and re forestation, or other antropogenic activities (urbanization, land reclamation) their numbers decrease, and they become more and more rare and endangered. During many years of investigations concerning the family Zygaenidae their primary and secondary parasitoids belonging to several families of Hymenoptera were reared. The host species were as follows: Adscita (Adscita) statices (L.), Zygaena (Mesembrynus) brizae (Esper), Z (Mesembrynus) minos (Denis & Schiffermuller), Z. -

Wildlife Review Cover Image: Hedgehog by Keith Kirk
Dumfries & Galloway Wildlife Review Cover Image: Hedgehog by Keith Kirk. Keith is a former Dumfries & Galloway Council ranger and now helps to run Nocturnal Wildlife Tours based in Castle Douglas. The tours use a specially prepared night tours vehicle, complete with external mounted thermal camera and internal viewing screens. Each participant also has their own state- of-the-art thermal imaging device to use for the duration of the tour. This allows participants to detect animals as small as rabbits at up to 300 metres away or get close enough to see Badgers and Roe Deer going about their nightly routine without them knowing you’re there. For further information visit www.wildlifetours.co.uk email [email protected] or telephone 07483 131791 Contributing photographers p2 Small White butterfly © Ian Findlay, p4 Colvend coast ©Mark Pollitt, p5 Bittersweet © northeastwildlife.co.uk, Wildflower grassland ©Mark Pollitt, p6 Oblong Woodsia planting © National Trust for Scotland, Oblong Woodsia © Chris Miles, p8 Birdwatching © castigatio/Shutterstock, p9 Hedgehog in grass © northeastwildlife.co.uk, Hedgehog in leaves © Mark Bridger/Shutterstock, Hedgehog dropping © northeastwildlife.co.uk, p10 Cetacean watch at Mull of Galloway © DGERC, p11 Common Carder Bee © Bob Fitzsimmons, p12 Black Grouse confrontation © Sergey Uryadnikov/Shutterstock, p13 Black Grouse male ©Sergey Uryadnikov/Shutterstock, Female Black Grouse in flight © northeastwildlife.co.uk, Common Pipistrelle bat © Steven Farhall/ Shutterstock, p14 White Ermine © Mark Pollitt, -

Land-Use Changes, Farm Management and the Decline of Butterflies Associated with Semi-Natural Grasslands in Southern Sweden
A peer-reviewed open-access journal Nature Conservation Land-use6: 31–48 (2013) changes, farm management and the decline of butterflies.... 31 doi: 10.3897/natureconservation.6.5205 APPLIED ECOLOGY http://www.pensoft.net/natureconservation Launched to accelerate biodiversity conservation Land-use changes, farm management and the decline of butterflies associated with semi-natural grasslands in southern Sweden Sven G. Nilsson1, Markus Franzén1,2, Lars B. Pettersson1,3 1 Biodiversity Unit, Department of Biology, Lund University, Ecology Building, SE-223 62 Lund, Sweden 2 UFZ Helmholtz Centre for Environmental Research, Department of Community Ecology, Theodor-Lieser- Straße 4, D-06120 Halle, Germany 3 Swedish Butterfly Monitoring Scheme, Lund University, Ecology Buil- ding, SE-223 62 Lund, Sweden Corresponding author: Lars B. Pettersson ([email protected]) Academic editor: L. Penev | Received 26 March 2013 | Accepted 30 October 2013 | Published 18 November 2013 Citation: Nilsson SG, Franzén M, Pettersson LB (2013) Land-use changes, farm management and the decline of butterflies associated with semi-natural grasslands in southern Sweden. Nature Conservation 18: 31–48. doi: 10.3897/ natureconservation.6.5205 Abstract Currently, we are experiencing biodiversity loss on different spatial scales. One of the best studied taxo- nomic groups in decline is the butterflies. Here, we review evidence for such declines using five systematic studies from southern Sweden that compare old butterfly surveys with the current situation. Additionally, we provide data on butterfly and burnet moth extinctions in the region’s counties. In some local areas, half of the butterfly fauna has been lost during the last 60–100 years. -

Fazekas OK.Qxd
Somogyi Múzeumok Közleményei 15: 145–156 Kaposvár, 2002 Adatok Magyarország Zygaenidae faunájának ismeretéhez (VII.) (Microlepidoptera: Zygaenidae) FAZEKAS IMRE FAZEKAS I.: Data to the knowledge of Hungary’s Zygaenidae (1) Mindenütt elterjedt és többnyire gyakori, euryök fauna, N º 7 . fajok. Abstract: The seventh part of a a presents the seven were (2) Mindenütt elterjedt, túlnyomóan közepes gyakori- collected in the Hungary. Having examined the Hungarian ságú fajok. collections author summarizes the results of his revisions about the Zygaenidae from different geographical regions of (3) Lokálisan elterjedt fajok, amelyek a megfelelõ élõ- Hungary. Taxonomic, zoogeographic and faunistic analysis of helyeken nagyobb számban is elõfordulhatnak. species presented is given. Jordanita fazekasi Efetov, 1998 is (4) Fõleg természetvédelmi területeken ismert fajok. a new record for Hungary. Figures of the genitalia and wing (5) Csak lokális populációk ismertek, fennmaradásuk pattern of several species which are difficult to determine are feltételezhetõ. given. (6) Elõfordulásuk gyakoriságának megállapítása a hi- ányos kutatások miatt bizonytalan. Bevezetés (7) Az 1960 elõtti elõfordulási adataiknak jó részét újabban nem sikerült igazolni. Lehetséges, hogy a A magyarországi Zygaenidae kutatásokat nevezék- faj regresszióban van. tani, taxonómiai és biológiai szempontból sokáig (8) Csak néhány izolált populáció ismert, a génkicse- GOZMÁNY (1963), majd FORSTER & WOHLFAHRT rélõdés lehetõsége bizonytalan, fennmaradásuk (1960) munkái határozták meg, s csak fokozatosan csak feltételezhetõ. nyert teret ALBERTI (1958), REISS & TREMEWAN (9) Csak 1–2 helyhez kötötten élõ, gyér számú popu- (1967) továbbá NAUMANN (1985) biológiai fajfogal- láció ismert. Rendszerint stenök fajok, fennmara- mon és flilogenetikai alapokon nyugvó szemlélete. Az dásuk bizonytalan. eltúlzott generikus felosztás illetve az intraspecifikus (10) 1960 óta eltûnt vagy feltételezhetõen kipusztult „taxonok” áttekinthetetlen „névzavarai” után, napjainkra populáció(k). -

ZYGAENIDAE 170 (4000) Five-Spot Burnet Zygaena Trifolii (Esper, 1783
18 19 ZYGAENIDAE Table 1. Summarised differences between Zygaena trifolii and Z. lonicerae latomarginata . 170 (4000) Five-spot Burnet Zygaena trifolii (Esper, 1783) ssp. decreta Verity, 1926 Local EXTERNAL Zygaena trifolii Zygaena trifolii Zygaena lonicerae ssp. palustrella Verity, 1926 Local CHARACTERS ssp. decreta ssp. palustrella ssp. latomarginata ssp. subsyracusia Verity, 1926 (Channel Islands) of adult 171 (3999) Narrow-bordered Five-spot Burnet Zygaena lonicerae (Scheven, 1777) ssp. latomarginata Tutt, 1899 Common Forewing Median pair of red spots often Median pair of red spots often Specimens with spots ssp. jocelynae Tremewan, 1962 RDB markings confluent; more extensive confluence confluent; more extensive confluence merged rare*. ssp. insularis Tremewan, 1960 (Ireland) of spots also frequent, including of spots also frequent, including specimens with all spots merged. specimens with all spots merged. Diagnostic external and ecological characters Hindwing Black terminal band generally Black terminal band generally Black terminal band generally The colonial nature of burnet moth populations and their consequently enhanced tendency to become markings broader (may require comparison broader (may require comparison narrower (may require comparison phenotypically distinct, has resulted in the raising of many sub-species. Even within these, more subtle variations of many examples). of many examples). of many examples). in appearance can occur between colonies of the same species. Forewing Apex less strongly pointed, more Apex less strongly pointed, more Apex more strongly pointed shape rounded (may require comparison rounded (may require comparison (may require comparison of In spite of much research, no clear and consistent differences in the genitalia, or other internal or external of many examples). of many examples). -

2017 Scottish Macro Moth List
SCOTTISH MACRO-MOTH LIST, 2017 Vernacular Name Code Taxon UK Status Scottish status Scottish Trend since 1980 Orange Swift 3.001 Triodia sylvina Common Widespread but local stable Common Swift 3.002 Korscheltellus lupulina Common Common S, scarce or absent N stable Map-winged Swift 3.003 Korscheltellus fusconebulosa Local Common stable Gold Swift 3.004 Phymatopus hecta Local Common stable Ghost Moth 3.005 Hepialus humuli Common Common stable Goat Moth 50.001 Cossus cossus Nb Scarce and very local, mainly Highlands stable? Lunar Hornet Moth 52.003 Sesia bembeciformis Common Widespread but overlooked? decline - G. S. Woodpecker predation? Welsh Clearwing 52.005 Synanthedon scoliaeformis RDB Very local in Highlands stable Large Red-belted Clearwing 52.007 Synanthedon culiciformis Nb Widespread but overlooked? stable Red-tipped Clearwing 52.008 Synanthedon formicaeformis Nb Dumfries & Galloway, last seen in 1942 extinct, or overlooked? Currant Clearwing 52.013 Synanthedon tipuliformis Nb SE only? Still present VC82 in 2014 major decline Thrift Clearwing 52.016 Pyropteron muscaeformis Nb SW & NE coasts, very local stable Forester 54.002 Adscita statices Local Very local, W and SW stable? Transparent Burnet 54.004 Zygaena purpuralis Na Very local, west Highlands stable Slender Scotch Burnet 54.005 Zygaena loti RDB Mull only stable Mountain Burnet 54.006 Zygaena exulans RDB Very local, VC92 stable New Forest Burnet 54.007 Zygaena viciae RDB, protected Very local, W coast fluctuates Six-spot Burnet 54.008 Zygaena filipendulae Common Common but mainly coastal range expansion NW, also inland Narrow-bordered Five-spot Burnet 54.009 Zygaena lonicerae Common SE, spreading; VC73; ssp. -
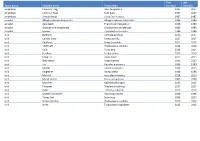
Taxon Group Common Name Taxon Name First Recorded Last
First Last Taxon group Common name Taxon name recorded recorded amphibian Common Frog Rana temporaria 1987 2017 amphibian Common Toad Bufo bufo 1987 2017 amphibian Smooth Newt Lissotriton vulgaris 1987 1987 annelid Alboglossiphonia heteroclita Alboglossiphonia heteroclita 1986 1986 annelid duck leech Theromyzon tessulatum 1986 1986 annelid Glossiphonia complanata Glossiphonia complanata 1986 1986 annelid leeches Erpobdella octoculata 1986 1986 bird Bullfinch Pyrrhula pyrrhula 2016 2017 bird Carrion Crow Corvus corone 2017 2017 bird Chaffinch Fringilla coelebs 2015 2017 bird Chiffchaff Phylloscopus collybita 2014 2016 bird Coot Fulica atra 2014 2014 bird Fieldfare Turdus pilaris 2015 2015 bird Great Tit Parus major 2015 2015 bird Grey Heron Ardea cinerea 2013 2017 bird Jay Garrulus glandarius 1999 1999 bird Kestrel Falco tinnunculus 1999 2015 bird Kingfisher Alcedo atthis 1986 1986 bird Mallard Anas platyrhynchos 2014 2015 bird Marsh Harrier Circus aeruginosus 2000 2000 bird Moorhen Gallinula chloropus 2015 2015 bird Pheasant Phasianus colchicus 2017 2017 bird Robin Erithacus rubecula 2017 2017 bird Spotted Flycatcher Muscicapa striata 1986 1986 bird Tawny Owl Strix aluco 2006 2015 bird Willow Warbler Phylloscopus trochilus 2015 2015 bird Wren Troglodytes troglodytes 2015 2015 bird Yellowhammer Emberiza citrinella 2000 2000 conifer Douglas Fir Pseudotsuga menziesii 2004 2004 conifer European Larch Larix decidua 2004 2004 conifer Lawson's Cypress Chamaecyparis lawsoniana 2004 2004 conifer Scots Pine Pinus sylvestris 1986 2004 crustacean -

Entomofauna Suppl Wgtremewan Z0.Indd 104 23.05.17 16:08 Entomofauna Suppl Wgtremewan Z0.Indd 105 23.05.17 16:08 W
This volume is dedicated to Gerry Tremewan, a famous Cornish entomologist who worked on Lepidoptera from the age of 12 until his death aged 85, specialising for over 60 years on the Zygaenidae. A foreword by his son Malcolm is followed by contributions from Emmanuelle Briolat, Roger L. H. Dennis, Eric Drouet, Konstantin A. Efetov, Axel Hofmann, Marc Nicolle, Adrian Spalding, Gerhard M. Tarmann, John R. G. Turner, Zoya Yefre- mova and Hossein Rajaei. The book lists his complete bibliography of 220 papers and books, all the 68 taxa Supplement 20, 224 Seiten ISSN 0250-4413 Ansfelden, 20. Juni 2017 described by him and the 7 taxa dedicated to him. This – 2016) (1931 special illustrated memorial book also includes Gerry’s own memoirs in a separate section entitled Have Net, Will Travel: Reminiscences of an old entomologist. Gerry Tremewan Gerry Tremewan Iran: Fars, Dasht-e Arzhan, 27.5.2013 Hofmann, Spalding, Tarmann & Witt (eds.) & Witt Hofmann, Spalding, Tarmann Axel Hofmann, Adrian Spalding, Gerhard Tarmann & Thomas Witt (eds.) Gerry Tremewan (1931 – 2016) The adventurous life of a Cornish entomologist Entomofauna_Suppl WGTremewan_02_US_02.indd 1-3 23.05.17 16:19 Gedruckt mit Unterstützung durch die Thomas-Witt-Stiftung zur Förderung der Wissenschaft und Forschung im Bereich der Zoologischen Systematik Entomofauna, Supplementum 20 ISSN 0250-4413 © Maximilian Schwarz, Ansfelden, 2017 Edited and Published by Maximilian Schwarz Konsulent f. Wissenschaft der Oberösterreichischen Landesregierung Eibenweg 6 4052 Ansfelden, Austria E-Mail: [email protected] -

Zygaenidae of Mongolia (Lepidoptera) 159-164 Nachr
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Nachrichten des Entomologischen Vereins Apollo Jahr/Year: 2011 Band/Volume: 32 Autor(en)/Author(s): Efetov Konstantin A., Gorbunov Oleg G., Tarmann Gerhard Michael Artikel/Article: Zygaenidae of Mongolia (Lepidoptera) 159-164 Nachr. entomol. Ver. Apollo, N. F. 32 (3/4): 159–164 (2012) 159 Zygaenidae of Mongolia (Lepidoptera) Konstantin A. Efetov, Oleg G. Gorbunov and Gerhard M. Tarmann Prof. Dr. Konstantin A. Efetov, Crimean State Medical University, UA-95006 Simferopol, Crimea, Ukraine; [email protected] Dr. Oleg G. Gorbunov, A. N. Severtsov Institute of Ecology and Evolution of Russian Academy of Sciences, Leninskiy prospekt, 33, RU-119071 Moscow, Russia; [email protected] Prof. Dr. Gerhard M. Tarmann, Tiroler Landesmuseen, Ferdinandeum, Naturwissenschaftliche Sammlungen, Feldstrasse 11a, A-6020 Innsbruck, Austria; [email protected] Abstract: An overview of the Zygaenidae fauna of Mongo - Zygaena (Agrumenia) exulans (Hohenwarth, 1792[: lia is provided. Ten species are recorded, including Il li be ris 265]) (Primilliberis) pruni Dyar, 1905, which is new for Mon go lia. (Sphinx sp.). Z. (A.) exulans sajana Die Zygaenidae der Mongolei (Lepidoptera) Burgeff, 1926(: 25). = altayensis Dabrowski, 1977(: 27). Zusammenfassung: Die derzeitige Kenntnis über die Zy gae- = chastilovi Churkin & Saldaitis, 2005(: 133). — Syn on y- ni dae-Arten der Mongolei wird zusammengefaßt. 10 Arten miz ed by Tremewan (2006: 144). werden aufgelistet. Eine Art wird erstmals für das Ge biet der Mongolei gemeldet: Illiberis (Primilliberis) pruni Dyar, 1905. Zygaena (Agrumenia) viciae ([Denis & Schiffermül- ler], 1775[: 45]) A checklist of the Zygaenidae of Mongolia (Sphinx sp.). -

ICN Pages June 09
Invertebrate Conservation News Number 66 October 2011 ISSN 1356 1359 Editor David Lonsdale A publication of The Amateur Entomologists' Society ntomol r E og u ist te s’ a S m o A c i e e t h y T Founded 1935 Where to write For all ICN business, please write to: AES P.O. Box 8774 London SW7 5ZG ICN Editor: David Lonsdale AES Habitat Conservation Officer: Vacant AES Conservation Committee Dafydd Lewis David Lonsdale Peter May Phil Wilkins ICN is printed by Cravitz Printing Co. Ltd., 1 Tower Hill, Brentwood, Essex CM14 4TA. Cover designed by Wayne Jarvis. INVERTEBRATE CONSERVATION NEWS No. 66, October 2011 EDITORIAL As pointed out six years ago in an ICN editorial (June 2005), losses of biodiversity cannot be prevented simply by the designation of sites that are considered to be of special importance for conservation. There is, however, little sign that this “inconvenient truth” has entered the minds of leading policy -makers, at least here in the UK. The protection of sites under various forms of official designation makes it possible to impose conservation measures that, for economic reasons, would generally not be feasible within the wider landscape. Within that wider landscape, however, a network of suitable sites is essential for the long-term survival of populations of many invertebrate species of limited mobility. If the successful dispersal of such species between sites is prevented, episodes of chance local extinction can lead to extinction at a regional, national or even global level. Also, where the geographic ranges of species are being affected by climate change, such dispersal is essential in the process whereby climate- induced retraction in part of a range can be matched by extension elsewhere. -

Xiv International Symposium on Zygaenidae 17-21 September 2014
XIV INTERNATIONAL SYMPOSIUM ON ZYGAENIDAE 17-21 SEPTEMBER 2014 TOBERMORY ISLE OF MULL SCOTLAND CONTENTS Contents & Sponsors 2 Welcome & Introduction 3 Participants 4 Programme 5-9 Abstracts 10-23 Front Page Photos: Slender Scotch burnet by kind permission of - Anand Prasad Transparent burnet by kind permission of - Tom Prescott Page 3 Header photo : Forester moth by kind permission of - Keith Tailby Page 11 Photo: Calgary Bay, Mull by kind permission of - Brigid Primrose THIS SYMPOSIUM IS SPONSORED BY 2 WELCOME & INTRODUCTION Welcome to the glorious island of Mull, one of Scotland’s Inner Hebridean islands and a haven for rare Zygaenidae. We do hope that you enjoy the Symposium but that equally you immerse yourself in the experience of staying in such an historic and picturesque place. The people of Mull are delighted to welcome visitors to their beautiful island and we will try and give you time to enjoy Tobermory and, on our excursions, we will include some of the wonderful scenery along the iconic coastline. Our Symposium includes papers on some Zygaenidae in very distant places; some of the results of recent work on the chemical ecology of Zygaenidae and the way that pheromones influence their lives; discussion of their ecology and behaviour; some recent work on the taxonomy of difficult species groups; and an account of conservation activities, focussing on Scottish burnet moths. Symposium papers will be presented at Glengorm Castle, north-west of Tobermory; the main outing will visit the north-west coast of Mull, before travelling down to the small island of Ulva; and then on Sunday 21st the papers focussed on Scottish species and their conservation will be at the Aros Hall in Tobermory. -

Annotated Host Catalogue for the Tachinidae (Diptera) of Italy
Stuttgarter Beiträge zur Naturkunde A, Neue Serie 3: 305–340; Stuttgart, 30.IV.2010. 305 Annotated host catalogue for the Tachinidae (Diptera) of Italy PIERFILIPPO CERRETTI & HANS-PETER TSCHORSNIG Abstract An annotated host catalogue is given for the Tachinidae of Italy. It comprises 180 tachinid species reared from 310 arthropod hosts belonging to seven insect orders and one chilopod order. The paper includes data from litera- ture as well as hitherto unpublished records. K e y w o r d s : Tachinidae, parasitoids, host catalogue, Italy. Zusammenfassung Für die italienischen Tachinidae wird ein kritischer Wirtekatalog gegeben. Er umfasst 180 Tachiniden-Arten und 310 Arthropoden-Wirtsarten aus sieben Insektenordnungen und einer Ordnung der Hundertfüßer. Die Arbeit enthält sowohl Daten aus der Literatur als auch bisher unveröffentlichte Wirtsangaben. Contents 1 Introduction .........................................................................................................................................................305 2 Explanation of the format ....................................................................................................................................306 3 Annotated parasitoid-host list .............................................................................................................................306 4 Host-parasitoid list ..............................................................................................................................................325 5 References ...........................................................................................................................................................330