Tation in the Winter and Dry Summers. Snowfall Varied from Slight to Trol Unit, a Clearcut Non-Burned Unit, and a Clearcut Slash
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pacific Water Shrew Sorex Bendirii
COSEWIC Assessment and Update Status Report on the Pacific Water Shrew Sorex bendirii in Canada ENDANGERED 2006 COSEWIC COSEPAC COMMITTEE ON THE STATUS OF COMITÉ SUR LA SITUATION ENDANGERED WILDLIFE DES ESPÈCES EN PÉRIL IN CANADA AU CANADA COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC 2006. COSEWIC assessment and update status report on the Pacific watershrew Sorex bendirii in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 28 pp. (www.sararegistry.gc.ca/status/status_e.cfm). Previous report: Galindo-Leal, C. and J.B. Runciman. 1994. COSEWIC status report on the Pacific water shrew Sorex bendirii in Canada. Committee on the Status of Endangered Wildlife in Canada. 1-33 pp. Production note: COSEWIC would like to acknowledge David Nagorsen for writing the update status report on the Pacific water shrew Sorex bendirii, prepared under contract with Environment Canada, and overseen and edited by Mark Brigham, Co-chair (Terrestrial Mammals), COSEWIC Terrestrial Mammals Species Specialist Subcommittee. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: (819) 997-4991 / (819) 953-3215 Fax: (819) 994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur la musaraigne de Bendire (Sorex bendirii) au Canada – Mise à jour. Cover illustration: Pacific water shrew – by Ron Altig. ©Her Majesty the Queen in Right of Canada 2006 Catalogue No. -

Tidal Marsh Recovery Plan Habitat Creation Or Enhancement Project Within 5 Miles of OAK
U.S. Fish & Wildlife Service Recovery Plan for Tidal Marsh Ecosystems of Northern and Central California California clapper rail Suaeda californica Cirsium hydrophilum Chloropyron molle Salt marsh harvest mouse (Rallus longirostris (California sea-blite) var. hydrophilum ssp. molle (Reithrodontomys obsoletus) (Suisun thistle) (soft bird’s-beak) raviventris) Volume II Appendices Tidal marsh at China Camp State Park. VII. APPENDICES Appendix A Species referred to in this recovery plan……………....…………………….3 Appendix B Recovery Priority Ranking System for Endangered and Threatened Species..........................................................................................................11 Appendix C Species of Concern or Regional Conservation Significance in Tidal Marsh Ecosystems of Northern and Central California….......................................13 Appendix D Agencies, organizations, and websites involved with tidal marsh Recovery.................................................................................................... 189 Appendix E Environmental contaminants in San Francisco Bay...................................193 Appendix F Population Persistence Modeling for Recovery Plan for Tidal Marsh Ecosystems of Northern and Central California with Intial Application to California clapper rail …............................................................................209 Appendix G Glossary……………......................................................................………229 Appendix H Summary of Major Public Comments and Service -

Terrestrial Mammal Species of Special Concern in California, Bolster, B.C., Ed., 1998 22
Terrestrial Mammal Species of Special Concern in California, Bolster, B.C., Ed., 1998 22 Salt marsh wandering shrew, Sorex vagrans halicoetes Paul W. Collins Description: A small to medium sized (100-110 mm TL), dark shrew, sooty seal brown to black above with a relatively long (37-41 mm), unicolored tail; dark brown ventrum; and moderately large high-domed skull (Grinnell 1913). Distinguished from upland S. v. vagrans by its darker dorsum and brown ventrum (silvery brown in vagrans), slightly larger body size, broader rostrum, and longer maxillary tooth-row (Grinnell 1913, Jackson 1928, Findley 1955). Distinguished from S. o. sinuosus by its slightly lighter color, larger size, browner ears, and high, dome-shaped cranium (Grinnell 1913). Weight from 3.1 to 7.2 g (males) and from 2.7 to 7.0 g (females) (Rudd 1955b). Distinguished from upland S. v. vagrans by darker dorsal and ventral pelage (Grinnell 1913). Taxonomic Remarks: The salt-marsh wandering shrew was first described as S. halicoetes Grinnell (1913). Jackson (1928) relegated halicoetes to a subspecies of S. vagrans, a convention followed by subsequent authors (Grinnell 1933, Findley 1955, Hennings and Hoffmann 1977, Junge and Hoffmann 1981, Carraway 1990). The taxonomy of S. vagrans group has had a confusing history (Merriam 1895, Grinnell 1913, Jackson 1928). There is little controversy about the taxonomic status of S. v. halicoetes, although the karyotype from halicoetes was identical to S. v. vagrans from the northern part of the San Francisco Bay region (Brown 1974). This taxon still needs a more thorough biochemical and morphometric analysis to help clarify its phylogenetic and taxonomic relationship to other members of the S. -

List of Species Included in ACE-II Native and Harvest Species Richness Counts (Appendix C)
Appendix C. Species included in native and harvest species richness counts. Animal Species Included in the Range Analysis....... ............................ ................................. C-01 Animal Species with Range Models Not Included in the Analysis......................... ................. C-20 Animal Species without Range Models, therefore not Included in the Analysis......... ............ C-20 Fish Species Included in the Range Analysis.................................................................... ....... C-30 Fish with Ranges not Included in the Analysis because neither Native nor Harvest................ C-33 Native Plants Species Included in the Range Analysis............................................................. C-34 CWHR Species and ACEII hexagon analysis of ranges. Of the 1045 species in the current CWHR species list used in ACE-II, 694 had range models digitized for use. Of those, 688 are included in this hexagon analysis of the state of Cailfornia, with 660 of those classified as native to California. There were no additional species picked up offshore due to the use of hexagon centroid points. Animal species INCLUDED in this analysis. CODE COMMON NAME SCIENTIFIC NAME A001 CALIFORNIA TIGER SALAMANDER Ambystoma californiense NATIVE A002 NORTHWESTERN SALAMANDER Ambystoma gracile NATIVE A003 LONG-TOED SALAMANDER Ambystoma macrodactylum NATIVE A047 EASTERN TIGER SALAMANDER Ambystoma tigrinum INTROD A021 CLOUDED SALAMANDER Aneides ferreus NATIVE A020 SPECKLED BLACK SALAMANDER Aneides flavipunctatus NATIVE A022 -

Revised Checklist of North American Mammals North of Mexico, 1986 J
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Mammalogy Papers: University of Nebraska State Museum, University of Nebraska State Museum 12-12-1986 Revised Checklist of North American Mammals North of Mexico, 1986 J. Knox Jones Jr. Texas Tech University Dilford C. Carter Texas Tech University Hugh H. Genoways University of Nebraska - Lincoln, [email protected] Robert S. Hoffmann University of Nebraska - Lincoln Dale W. Rice National Museum of Natural History See next page for additional authors Follow this and additional works at: http://digitalcommons.unl.edu/museummammalogy Part of the Biodiversity Commons, Other Ecology and Evolutionary Biology Commons, Terrestrial and Aquatic Ecology Commons, and the Zoology Commons Jones, J. Knox Jr.; Carter, Dilford C.; Genoways, Hugh H.; Hoffmann, Robert S.; Rice, Dale W.; and Jones, Clyde, "Revised Checklist of North American Mammals North of Mexico, 1986" (1986). Mammalogy Papers: University of Nebraska State Museum. 266. http://digitalcommons.unl.edu/museummammalogy/266 This Article is brought to you for free and open access by the Museum, University of Nebraska State at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Mammalogy Papers: University of Nebraska State Museum by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Authors J. Knox Jones Jr., Dilford C. Carter, Hugh H. Genoways, Robert S. Hoffmann, Dale W. Rice, and Clyde Jones This article is available at DigitalCommons@University of Nebraska - Lincoln: http://digitalcommons.unl.edu/museummammalogy/ 266 Jones, Carter, Genoways, Hoffmann, Rice & Jones, Occasional Papers of the Museum of Texas Tech University (December 12, 1986) number 107. U.S. -

Chapter 7 Small Mammal Distribution, Abundance and Habitat Use
CHAPTER 7 SMALL MAMMAL DISTRIBUTION, ABUNDANCE AND HABITAT USE by Klaus O. Richter and Amanda L. Azous INTRODUCTION Small mammals are an integral component of most ecosystems. In the Northwest the regional distribution of small mammals has been described by Ingles (1965) and Maser et al. (1981). Within unmanaged (e.g., old growth) Douglas-fir forests, small mammals were described in Aubry et al. (1991) by numerous biologists. Mammals in second growth forests under differing cutting practices and intensity of landscaping and development were described by (Stofel 1993). Local small mammal species and distributions in urban parks varying in size of approximately four to 400 ha within urbanizing areas were described by (Gavareski 1976). The distribution and abundance of small mammals, similar to that of macroinvertebrates, amphibians and birds may be indicators of the environmental health of wetlands. They also exhibit the ability to shape wetlands through their influence on soil, water and plants. Several species such as Trowbridge’s shrew, marsh shrew, shrew-mole, western red-backed vole and creeping vole (See Table 7-1 for scientific names) are endemic to the Pacific Northwest (Corn and Bury 1991) and could be expected at pristine wetlands. Our objective in this chapter is to present the relative distribution and abundance of small mammals across the wetlands we studied. We also examine wetland conditions such as size, hydrology and vegetation complexity to gain insight into habitat characteristics important for maintaining diversity and unique species. METHODS We used pitfall and Sherman trap captures during autumn (mid-October to mid- November) as indicators of small mammal distributions. -

Life History Account for Vagrant Shrew
California Wildlife Habitat Relationships System California Department of Fish and Wildlife California Interagency Wildlife Task Group VAGRANT SHREW Sorex vagrans Family: SORICIDAE Order: INSECTIVORA Class: MAMMALIA M003 Written by: J. Harris Reviewed by: H. Shellhammer Edited by: S. Granholm, R. Duke Updated by: CWHR Program Staff, May 2000 DISTRIBUTION, ABUNDANCE, AND SEASONALITY The vagrant shrew is common to abundant in the Sierra Nevada and Cascades from the Oregon border to northern Inyo Co., and along the coast from the Oregon border to Monterey Bay. Optimal habitats are valley foothill and montane riparian, aspen, wet meadow, annual and perennial grasslands, and fresh and saline emergent wetlands. Also occurs in a variety of chaparral and wooded habitats. Vagrant shrews can be found in California from sea level to 3750 m (12,000 ft) (Findley 1999). SPECIFIC HABITAT REQUIREMENTS Feeding: Feeds mainly on invertebrates: insects, worms, snails, slugs, and spiders. Also eats fungi, small mammals, roots, young shoots, and probably seeds. Forages under litter on moist surfaces, underground, and in moist accumulations of dead plant material. Whitaker et al. (1983) reported it ate more flying insects and caterpillars, and fewer terrestrial insects and worms, when grazing pressure was heavy. Cover: Prefers dense litter or ground cover. Uses vole runways. Reproduction: Makes a nest of dry grass, moss, or other materials under logs, roots, or dense vegetation. (Findley 1999). Water: This species may require water. Clothier (1955) reported that vagrant shrews were not found farther than 180 m (600 ft) from water. Pattern: Prefers patchy, open areas with wet micro-habitats such as damp meadows and streambanks. -

Mammals and Life Zones
Mount Rainier National Park National Park Service U.S. Department of the Interior Mammals and Life Zones Just as people have adapted to living in nearly every climate imaginable, our fellow mammals have come to occupy many different life zones around the world. These life zones, like big neighborhoods, provide many different habitats where animals can find food, water, shelter, and space. More than fifty different kinds of mammals live in the life zones found in Mount Rainier National Park. Some are specially adapted to one life zone, while others range through several. Imagine taking a very long walk from the edge of the park up to the summit of the mountain. What kinds of life zones would you see? What kinds of mammals would you find in each one? Looking at life zones can help us better understand and appreciate mammal adaptations and the struggle to survive––a trait all living things share. Mountain goats live in the Alpine Zone Lowland Forest Zone You’re in the lowland forest when you enter the park and find yourself sheltered by giant trees, dense shrubs and brush. The old-growth forests of this zone have trees towering 250 feet (76 m) or more, reaching diameters of 100 inches (2.54 m). Found between 2,000 and 3,000 feet in elevation (610-914 m), this zone is crowded with Douglas fir, western hemlock, and western red cedar. If you pass a pond or a lake, you might see evidence of beavers. Beavers eat tree bark and “girdle” trees which they can later use to make dams and lodges. -

Complete List of Amphibian, Reptile, Bird and Mammal Species in California
Complete List of Amphibian, Reptile, Bird and Mammal Species in California California Department of Fish and Game Sept. 2008 (updated) This list represents all of the native or introduced amphibian, reptile, bird and mammal species known in California. Introduced species are marked with “I”, harvest species with “HA”, and vagrant species or species with extremely limited distributions with *. The term “introduced”, as used here, represents both accidental and intentional introductions. Subspecies are not included on this list. The most current list of species and subspecies with special management status is available from the California Natural Diversity Database (CNDDB) Taxonomy and nomenclature used within the list are the same as those used within both the CNDDB and CWHR software programs and data sets. If a discrepancy exists between this list and the ones produced by CNDDB, the CNDDB list can be presumed to be more accurate as it is updated more frequently than the CWHR data set. ________________________________________________________________________ ______________________________________________________________________ ______________________________________________________________________ AMPHIBIA (Amphibians) CAUDATA (Salamanders) AMBYSTOMATIDAE (Mole Salamanders and Relatives) Long-toed Salamander Ambystoma macrodactylum Tiger Salamander Ambystoma tigrinum I California Tiger Salamander Ambystoma californiense Northwestern Salamander Ambystoma gracile RHYACOTRITONIDAE (Torrent or Seep Salamanders) Southern Torrent Salamander Rhyacotriton -

SHREWS Robert H
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln The aH ndbook: Prevention and Control of Wildlife Wildlife Damage Management, Internet Center for Damage 1-1-1994 SHREWS Robert H. Schmidt Utah State University, [email protected] Follow this and additional works at: http://digitalcommons.unl.edu/icwdmhandbook Part of the Environmental Sciences Commons Schmidt, Robert H., "SHREWS" (1994). The Handbook: Prevention and Control of Wildlife Damage. 55. http://digitalcommons.unl.edu/icwdmhandbook/55 This Article is brought to you for free and open access by the Wildlife Damage Management, Internet Center for at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in The aH ndbook: Prevention and Control of Wildlife Damage by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Robert H. Schmidt Assistant Professor Department of Fisheries and Wildlife SHREWS Utah State University Logan, Utah 84322-5210 Fig. 1. A masked shrew, Sorex cinereus Identification Damage Prevention and Fumigants The shrew is a small, mouse-sized Control Methods None are registered. mammal with an elongated snout, a Trapping dense fur of uniform color, small eyes, Exclusion and five clawed toes on each foot (Fig. Mouse trap (snap trap). Rodent-proof structures also exclude 1). Its skull, compared to that of shrews. Small box trap. rodents, is long, narrow, and lacks the zygomatic arch on the lateral side Cultural Methods Pit trap. characteristic of rodents. The teeth are Mowing may decrease preferred Shooting small, sharp, and commonly dark- tipped. Pigmentation on the tips of the habitat and food. Not practical. teeth is caused by deposition of iron in Repellents Other Methods the outer enamel. -

Diversity and Endemism in Tidal-Marsh Vertebrates
Studies in Avian Biology No. 32:32-53 DIVERSITY AND ENDEMISM IN TIDAL-MARSH VERTEBRATES RUSSELL GREENBERG AND JESúS E. MALDONADO Abstract. Tidal marshes are distributed patcliily, predominantly along the mid- to high-latitude coasts of the major continents. The greatest extensions of non-arctic tidal marshes are found along the Atlantic and Gulf coasts of North America, but local concentrations can be found in Great Britain, northern Europe, northern Japan, northern China, and northern Korea, Argentina-Uruguay-Brazil, Australia, and New Zealand. We tallied the number of terrestrial vertebrate species that regularly occupy tidal marshes in each of these regions, as well as species or subspecies that are largely restricted to tidal marshes. In each of the major coastal areas we found 8-21 species of breeding birds and 13-25 species of terrestrial mammals. The diversity of tidal-marsh birds and mammals is highly inter-correlated, as is the diversity of species restricted to saltmarshes. These values are, in turn, correlated with tidal-marsh area along a coastline. We estimate approximately seven species of turtles occur in brackish or saltmarshes worldwide, but only one species is endemic and it is found in eastern North America. A large number of frogs and snakes occur opportunistically in tidal marshes, primarily in southeastern United States, particularly Florida. Three endemic snake taxa are restricted to tidal marshes of eastern North America as well. Overall, only in North America were we able to find documentation for multiple taxa of terrestrial vertebrates associated with tidal marshes. These include one species of mammal and two species of birds, one species of snake, and one species of turtle. -
Ef-Funa Mammal Checklist 2014 Version
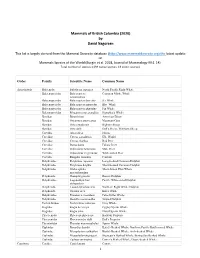
Mammals of British Columbia (2020) by David Nagorsen This list is largely derived from the Mammal Diversity database (http://www.mammaldiversity.org) the latest update on Mammals Species of the World (Burgin et al. 2018, Journal of Mammalogy 99:1-14) . Total number of species: (139 native species; 13 exotic species). Order Family Scientific Name Common Name Artiodactyla Balaenidae Eubalaena japonica North Pacific Right Whale Balaenopteridae Balaenoptera Common Minke Whale acutorostrata Balaenopteridae Balaenoptera borealis Sei Whale Balaenopteridae Balaenoptera musculus Blue Whale Balaenopteridae Balaenoptera physalus Fin Whale Balaenopteridae Megaptera novaeangliae Humpback Whale Bovidae Bison bison American Bison Bovidae Oreamnos americanus Mountain Goat Bovidae Ovis canadensis Bighorn Sheep Bovidae Ovis dalli Dall's Sheep, Thinhorn Sheep Cervidae Alces alces Moose Cervidae Cervus canadensis Elk, Wapiti Cervidae Cervus elaphus Red Deer Cervidae Dama dama Fallow Deer Cervidae Odocoileus hemionus Mule Deer Cervidae Odocoileus virginianus White‐tailed Deer Cervidae Rangifer tarandus Caribou Delphinidae Delphinus capensis Long‐beaked Common Dolphin Delphinidae Delphinus delphis Short‐beaked Common Dolphin Delphinidae Globicephala Short‐finned Pilot Whale macrorhynchus Delphinidae Grampus griseus Risso's Dolphin Delphinidae Lagenorhynchus Pacific White‐sided Dolphin obliquidens Delphinidae Lissodelphis borealis Northern Right Whale Dolphin Delphinidae Orcinus orca Killer Whale Delphinidae Pseudorca crassidens False Killer Whale Delphinidae