New Species of Springtails in the Proisotoma Genus Complex From
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Folsomia Candida and the Results of a Ringtest
Toxicity testing with the collembolans Folsomia fimetaria and Folsomia candida and the results of a ringtest P.H. Krogh DMU/AU, Denmark Department of Terrestrial Ecology With contributions from: Mónica João de Barros Amorim, Pilar Andrés, Gabor Bakonyi, Kristin Becker van Slooten, Xavier Domene, Ine Geujin, Nobuhiro Kaneko, Silvio Knäbe, Vladimír Kocí, Jan Lana, Thomas Moser, Juliska Princz, Maike Schaefer, Janeck J. Scott-Fordsmand, Hege Stubberud, Berndt-Michael Wilke August 2008 1 Contents 1 PREFACE 3 2 BIOLOGY AND ECOTOXICOLOGY OF F. FIMETARIA AND F. CANDIDA 4 2.1 INTRODUCTION TO F. FIMETARIA AND F. CANDIDA 4 2.2 COMPARISON OF THE TWO SPECIES 6 2.3 GENETIC VARIABILITY 7 2.4 ALTERNATIVE COLLEMBOLAN TEST SPECIES 8 2.5 DIFFERENCES IN SUSCEPTIBILITY OF THE TWO SPECIES 8 2.6 VARIABILITY IN REPRODUCTION RATES 8 3 TESTING RESULTS OBTAINED AT NERI, 1994 TO 1999 10 3.1 INTRODUCTION 10 3.2 PERFORMANCE 10 3.3 INFLUENCE OF SOIL TYPE 10 3.4 CONCLUSION 11 4 RINGTEST RESULTS 13 4.1 TEST GUIDELINE 13 4.2 PARTICIPANTS 13 4.3 MODEL CHEMICALS 14 4.4 RANGE FINDING 14 4.5 STATISTICAL ANALYSIS 14 4.6 EXPERIMENTAL DESIGN 15 4.7 TEST CONDITIONS 15 4.8 CONTROL MORTALITY 15 4.9 CONTROL REPRODUCTION 16 4.10 VARIABILITY OF TESTING RESULTS 17 4.11 CONCLUSION 18 5 SUMMARY AND CONCLUSIONS 27 6 ACKNOWLEDGEMENTS 29 7 REFERENCES 30 ANNEX 1 PARTICIPANTS 36 ANNEX 2 LABORATORY CODE 38 ANNEX 3 BIBLIOMETRIC STATISTICS 39 ANNEX 4 INTRALABORATORY VARIABILITY 40 ANNEX 5 CONTROL MORTALITY AND REPRODUCTION 42 ANNEX 6 DRAFT TEST GUIDELINE 44 2 1 Preface Collembolans have been used for ecotoxicological testing for about 4 decades now but they have not yet had the privilege to enter into the OECD test guideline programme. -

Grand Isle State Park
Click on a Site for Photo and Additional Information CYPRESS SHAD WILLOW Grand Isle State Park FORESTS, PARKS & RECREATION APPLE VERMONT FIR Grand Isle, Vermont ASH AGENCY OF NATURAL RESOURCES ELM HEMLOCK OAK CEDAR 1A North BEECH V 1 2 MAPLE 3 4 2 5 PINE 6 7 9 8 29 27 STERLING 25 23 10 21 20 4 19 11 12 JUNIPER 18 17 47 46 28 26 24 15 Lake Champlain 30 22 13 SYCAMORE 32 34 36 38 44 16 14 CHESTNUT 40 42 48 31 45 HAWTHORN 33 35 37 49 39 BUTTERNUT 78 77 76 41 43 BIRCH 74 72 70 68 YEW LARCH 1 66 71 CAMELS 75 73 69 BOLTON 67 MANSFIELD HUMP SPRUCE 65 110 MTN POPLAR BALSAM 112 MULBERRY BOXELDER V 64 53 55 57 59 61 102 DOGWOOD CHERRY WALNUT 63 111 108 106 ALDER SUMAC 113 54 3 103 101 56 58 6 119 LOCUST 60 62 115 V 121 123 80 BASSWOOD 117 118 120 TAMARACK 114 122 HICKORY ASPEN 84 99 88 90 92 94 PROPERTY 81 86 96 100 PRIVATE 82 83 98 Park office Drinking water 85 87 89 91 93 95 97 0 Prime campsite Trash/Recycling to Grand Isle Village East Shore South 0 Campsite V Volunteer Site park entrance 0 100 200 400 600 to Prime lean-to Parking 2 feet this map is intended for informational purposes only Lean-to Nature Center Wheelchair Playground Cabin accessible facility Horseshoe pit Pets allowed on leash Restrooms RV Sanitary Station on trail Steep bank/ Cliffs Volleyball Trail Showers ($) Boat launch Traffic direction Swimming area ephelps-rev. -

AND WESTERN SYRIA PART III FAMILY ISOTOMIDAE the Work
THE COLLEMBOLA OF LEBANON AND WESTERN SYRIA PART III FAMILY ISOTOMIDAE BY K. CHRISTIANSEN Grinnell College, Grinnell, Iowa The work herein described was done under N.S.F. Grant G 4563. I wish to gratefully acknowledge the work of my assistant, Jerry Tecklin, who did all of the preliminary sorting and most of the handling and mounting of the material here studied. Relatively little has been known about the Isotomidae of the Syrian region. Until the work of Cassagnau and Delamare only a few of the more prominent epigeic forms had been described. With the above mentioned work th.e recorded species from the area were eight. In the present study four of these forms were recovered and two others were probably recovered. In addition 16 new records were established. Anurophorus coiffaiti Cassagnau & Delamare Plate 8, figures 6, 7 Anurophorus coi]]aii P. Cassagnau & C1. Delamore, 1951 Biospeologic 75:377,378. This species was the first of the genus to be described from Western Syria. The specimens at hand agree well with the illustrations and figures given by Delamare and Cassagnau. The P.A.o. (see figure 7) is characteristically oval with a definite indentation or indication of a listel at least on the forward margin. The abdominal thickenings chararacteristic of the species vary a great deal but some indication of these can be seen even in the youngest speci- mens. In some forms these fuse into a single large ubercle- like projection. The sense organ of the third antennal segment usually has only two short setae between the Published with the aid of a grant from the Museum of Comparative Zoology at Harvard College. -

Curt Teich Postcard Archives Towns and Cities
Curt Teich Postcard Archives Towns and Cities Alaska Aialik Bay Alaska Highway Alcan Highway Anchorage Arctic Auk Lake Cape Prince of Wales Castle Rock Chilkoot Pass Columbia Glacier Cook Inlet Copper River Cordova Curry Dawson Denali Denali National Park Eagle Fairbanks Five Finger Rapids Gastineau Channel Glacier Bay Glenn Highway Haines Harding Gateway Homer Hoonah Hurricane Gulch Inland Passage Inside Passage Isabel Pass Juneau Katmai National Monument Kenai Kenai Lake Kenai Peninsula Kenai River Kechikan Ketchikan Creek Kodiak Kodiak Island Kotzebue Lake Atlin Lake Bennett Latouche Lynn Canal Matanuska Valley McKinley Park Mendenhall Glacier Miles Canyon Montgomery Mount Blackburn Mount Dewey Mount McKinley Mount McKinley Park Mount O’Neal Mount Sanford Muir Glacier Nome North Slope Noyes Island Nushagak Opelika Palmer Petersburg Pribilof Island Resurrection Bay Richardson Highway Rocy Point St. Michael Sawtooth Mountain Sentinal Island Seward Sitka Sitka National Park Skagway Southeastern Alaska Stikine Rier Sulzer Summit Swift Current Taku Glacier Taku Inlet Taku Lodge Tanana Tanana River Tok Tunnel Mountain Valdez White Pass Whitehorse Wrangell Wrangell Narrow Yukon Yukon River General Views—no specific location Alabama Albany Albertville Alexander City Andalusia Anniston Ashford Athens Attalla Auburn Batesville Bessemer Birmingham Blue Lake Blue Springs Boaz Bobler’s Creek Boyles Brewton Bridgeport Camden Camp Hill Camp Rucker Carbon Hill Castleberry Centerville Centre Chapman Chattahoochee Valley Cheaha State Park Choctaw County -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

Southeast Region
VT Dept. of Forests, Parks and Recreation Mud Season Trail Status List is updated weekly. Please visit www.trailfinder.info for more information. Southeast Region Trail Name Parcel Trail Status Bear Hill Trail Allis State Park Closed Amity Pond Trail Amity Pond Natural Area Closed Echo Lake Vista Trail Camp Plymouth State Park Caution Curtis Hollow Road Coolidge State Forest (east) Open Slack Hill Trail Coolidge State Park Closed CCC Trail Coolidge State Park Closed Myron Dutton Trail Dutton Pines State Park Open Sunset Trail Fort Dummer State Park Open Broad Brook Trail Fort Dummer State Park Open Sunrise Trail Fort Dummer State Park Open Kent Brook Trail Gifford Woods State Park Closed Appalachian Trail Gifford Woods State Park Closed Old Growth Interpretive Trail Gifford Woods State Park Closed West River Trail Jamaica State Park Open Overlook Trail Jamaica State Park Closed Hamilton Falls Trail Jamaica State Park Closed Lowell Lake Trail Lowell Lake State Park Closed Gated Road Molly Beattie State Forest Closed Mt. Olga Trail Molly Stark State Park Closed Weathersfield Trail Mt. Ascutney State Park Closed Windsor Trail Mt. Ascutney State Park Closed Futures Trail Mt. Ascutney State Park Closed Mt. Ascutney Parkway Mt. Ascutney State Park Open Brownsville Trail Mt. Ascutney State Park Closed Gated Roads Muckross State Park Open Healdville Trail Okemo State Forest Closed Government Road Okemo State Forest Closed Mountain Road Okemo State Forest Closed Gated Roads Proctor Piper State Forest Open Quechee Gorge Trail Quechee Gorge State Park Caution VINS Nature Center Trail Quechee Gorge State Park Open Park Roads Silver Lake State Park Open Sweet Pond Trail Sweet Pond State Park Open Thetford Academy Trail Thetford Hill State Park Closed Gated Roads Thetford Hill State Park Open Bald Mt. -

New and Little Known Isotomidae (Collembola) from the Shore of Lake Baikal and Saline Lakes of Continental Asia
ZooKeys 935: 1–24 (2020) A peer-reviewed open-access journal doi: 10.3897/zookeys.935.49363 RESEARCH ARTICLE https://zookeys.pensoft.net Launched to accelerate biodiversity research New and little known Isotomidae (Collembola) from the shore of Lake Baikal and saline lakes of continental Asia Mikhail Potapov1,2, Cheng-Wang Huang3, Ayuna Gulgenova4, Yun-Xia Luan5 1 Senckenberg Museum of Natural History Görlitz, Am Museum 1, 02826 Görlitz, Germany 2 Moscow Pedagogical State University, Moscow, 129164, Kibalchicha St. 6 b. 5, Russia 3 Key Laboratory of Insect Devel- opmental and Evolutionary Biology, CAS Center for Excellence in Molecular Plant Sciences, Chinese Academy of Sciences, Shanghai, 200032, China 4 Banzarov Buryat State University, Ulan-Ude, 670000, Smolina St. 24a, Russia 5 Guangdong Provincial Key Laboratory of Insect Developmental Biology and Applied Technology, Institute of Insect Science and Technology, School of Life Sciences, South China Normal University, Guangzhou, 510631, China Corresponding author: Cheng-Wang Huang ([email protected]) Academic editor: Wanda M. Weiner | Received 13 December 2019 | Accepted 13 March 2020 | Published 21 May 2020 http://zoobank.org/69778FE4-EAD8-4F5D-8F73-B8D666C25546 Citation: Potapov M, Huang C-W, Gulgenova A, Luan Y-X (2020) New and little known Isotomidae (Collembola) from the shore of Lake Baikal and saline lakes of continental Asia. ZooKeys 935: 1–24. https://doi.org/10.3897/ zookeys.935.49363 Abstract Collembola of the family Isotomidae from the shores of Lake Baikal and from six saline lake catenas of the Buryat Republic (Russia) and Inner Mongolia Province (China) were studied. Pseudanurophorus barathrum Potapov & Gulgenova, sp. -

Appendix a Places to Visit and Natural Communities to See There
Appendix A Places to Visit and Natural Communities to See There his list of places to visit is arranged by biophysical region. Within biophysical regions, the places are listed more or less north-to-south and by county. This list T includes all the places to visit that are mentioned in the natural community profiles, plus several more to round out an exploration of each biophysical region. The list of natural communities at each site is not exhaustive; only the communities that are especially well-expressed at that site are listed. Most of the natural communities listed are easily accessible at the site, though only rarely will they be indicated on trail maps or brochures. You, the naturalist, will need to do the sleuthing to find out where they are. Use topographic maps and aerial photographs if you can get them. In a few cases you will need to do some serious bushwhacking to find the communities listed. Bring your map and compass, and enjoy! Champlain Valley Franklin County Highgate State Park, Highgate Vermont Department of Forests, Parks, and Recreation Temperate Calcareous Cliff Rock River Wildlife Management Area, Highgate Vermont Department of Fish and Wildlife Silver Maple-Sensitive Fern Riverine Floodplain Forest Alder Swamp Missisquoi River Delta, Swanton and Highgate Missisquoi National Wildlife Refuge U.S. Fish and Wildlife Service Protected with the assistance of The Nature Conservancy Silver Maple-Sensitive Fern Riverine Floodplain Forest Lakeside Floodplain Forest Red or Silver Maple-Green Ash Swamp Pitch Pine Woodland Bog -
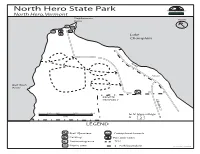
North Hero Map and Guide
North Hero State Park FORESTS, PARKS & RECREATION VERMONT North Hero, Vermont AGENCY OF NATURAL RESOURCES Stephenson North Point Lake Champlain PRIVATE PRIVATE Bull Rush Point PRIVATE PROPERTY Lakeview Dr. 0 150 300 600 900 to N. Hero village feet & 2 LEGEND Staff Quarters Cartop boat launch Parking Portable toilet Swimming area Trail Picnic area Park boundary ephelps-revised 03/2019 Isle LaMotte North Hero State Park ● St. Anne’s Shrine ● Ancient coral reef Welcome to North Hero State Park. Land for North Hero this 399-acre park was purchased in 1963. North Hero Nearly one-third of the land area lies below 100 ●Knight Point State Park feet in elevation. Lake Champlain normally State Park fluctuates from about 95 to 101 feet above sea Milton level, subjecting much of the park to seasonal ● Sand Bar State Park Map & Guide inundation. The forest type in the floodplain area is uncommon in Vermont, found only around Alburgh Lake Champlain. The lakeside floodplain forest ● Alburgh Dunes State Park at North Hero is noted for its size, relatively ● Lake Champlain Bikeways undisturbed condition and the valuable wildlife habitat it provides. For More Information contact: Wildlife habitat improvements at North Hero North Hero State Park State Park have yielded tangible results. White- 3803 Lakeview Drive tailed deer are common, as are a variety of North Hero, VT 05474 migratory waterfowl - mallards, black and wood (802) 372-8727 (Operating Season) ducks nest in the wooded wetlands. Ruffed Or Call grouse and American woodcock find suitable VT State Parks Reservations Center breeding and nesting habitat here as well. -

Brief Introduction Camp Plymouth State Park
State Parks In Vermont: Brief Introduction by newsdesk Camp Plymouth State Park :The site of Camp Plymouth was at one time thought to have been used as an encampment by soldiers of the Revolutionary War in 1777. The Boy Scouts used this area until 1984 when it became a state park. Camp Plymouth State Park is located in the town of Plymouth on the east shore of Echo Lake. The total acreage is 295 acres of which 46 acres comprise the developed portion of the park. The balance (249 acres) contains hiking trails, fishing, hunting, gold panning, and primitive camping, but is largely forestry oriented. Fort Dummer State Park :The park was named after Fort Dummer, the first permanent white settlement in Vermont. Built on the frontier in 1724, it was initially the gateway to the early settlements along the banks of the Connecticut River. Forty-three English soldiers and twelve Mohawk Indians manned the fort in 1724 and 1725. Later, the fort protected what was then a Massachusetts colony from an invasion by the French and Indians. Made of sturdy white pine timber, stacked like a log cabin, Fort Dummer served its purpose well. The park overlooks the site of Fort Dummer which was flooded when the Vernon Dam was built on the Connecticut River in 1908. This site can be seen from the northernmost scenic vista on the Sunrise Trail. It is now underwater near the lumber company located on the western bank of the river. Knight Point State Park :Knight Point on North Hero Island opened as a state park in 1978. -

Cyanobacteria Monitoring on Lake Champlain Summer 2013
Cyanobacteria Monitoring on Lake Champlain Summer 2013 Final Report for the Lake Champlain Basin Program May 2014 Prepared by Angela Shambaugh Watershed Management Division Vermont Agency of Natural Resources Montpelier, Vermont in conjunction with Andy Chevrefils Mike Winslow Radiation and Toxicological Section Lake Champlain Committee Vermont Department of Health Burlington, VT 1 Executive Summary Cyanobacteria monitoring on Lake Champlain in 2013 continued to integrate qualitative observations, photographic documentation, quantitative analysis of algae populations, and microcystin concentrations into guidance for Lake Champlain users. Additional monitoring on four Vermont lakes (Carmi, Elmore, Iroquois and Memphremagog) was made possible by a CDC Climate Change grant awarded to the Vermont Department of Health. Objectives continue to monitor cyanobacteria at locations on Lake Champlain through the established partnership between state and local officials, the Lake Champlain Committee and citizen volunteers; continue to provide consistent quantitative data at selected locations around Lake Champlain; test for the presence of microcystin and anatoxin when algal density and composition triggers are reached; facilitate communication about lake conditions through weekly updates to stakeholders via email and to the general public through the Vermont Department of Health webpage; continue to provide outreach and assistance to beach managers, lakeshore property owners and the general public so they can learn to recognize and respond appropriately to the presence of cyanobacteria blooms More than 800 site-specific reports were submitted during 2013 from 83 locations on Lake Champlain and the four inland lakes. Fifty-four Champlain locations were monitored by citizen volunteers trained by the Lake Champlain Committee. Blooms, defined as category 3 of the visual protocol and alert level 2 of the tiered alert protocol, were reported 14 times in 2013. -

IMPACT of INSECTICIDES on ABUNDANCE of NON-TARGET SOIL MESOFAUNA in RICE ECOSYSTEM E:\ 82(1) 19167-Ingle Dipak Shyamrao 2020
E:\ 82(1) 19167-Ingle Dipak Shyamrao 2020 Indian Journal of Entomology, 82(1): 000-000 (2020) DoI No.: IMPACT OF INSECTICIDES ON ABUNDANCE OF NON-TARGET SOIL MESOFAUNA IN RICE ECOSYSTEM INGLE DIPAK SHYAMRAO*, M. RAGHURAMAN, ABHINAV KUMAR AND RUPESH KUMAR GAJBHIYE Department of Entomology and Agricultural Zoology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi 221005 *Email: [email protected] (corresponding author) ABSTRACT An experimental trial was conducted during kharif 2016 and 2017 at the Agriculture Research Farm, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi to study the biodiversity and impact of conventional and newer insecticides on non-target soil mesofauna (springtails) in rice ecosystem. Result revealed that a total of 21 specimens under 14 genera of Collembola belonging to 7 families were studied from rice ecosystems. The species viz., Lepidcyrtus fimetarius, L. curvicollis, L. paradoxus, Proisotoma ripicola, Hypogastrura sonapani, H. viatica, Salina selebensis, Isotoma dagamae and I. trispinata were highly abundant. All the insecticidal treatments showed adverse effects on the collembolan population in the rice ecosystem. Considering all the four observations taken after the second insecticidal spray, neem (azadirachtin 0.15% EC) @ 4 ml/l followed by dinotefuran 20% SG @ 40 g a.i./ ha was found to be less detrimental. Fipronil 5% SC @ 50 g a.i./ ha resulted in drastic reduction in collembolan population followed by carbofuran 3% G @ 750 g a.i./ ha. Maximum reduction was observed with fipronil 5% SC @ 50 g a.i./ha with 57.14 and 56.00% during kharif, 2016 and 2017. Key words: Rice, insecticides, Collembola, azadirachtin, dinotefuran, fipronil, carbofuran, non-m target effects Rice is the main food and major cereal crop in India MATERIALS AND METHODS with a productivity of 3.78 t/ ha (Anonymous, 2016; Saxena and Singh, 2003).