COQMON-7 | Lower Coquitlam River Fish Productivity Index
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
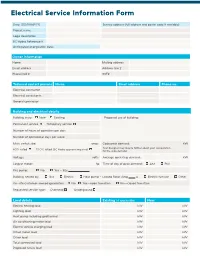
Electrical Service Information Form
Electrical Service Information Form Date: (DD/MM/YY) Service address (full address and postal code if available) Project name Legal description BC Hydro Reference # Anticipated energization date: Owner information Name Mailing address Email address Address line 2 Phone/Cell # GST# Technical contact persons Name Email address Phone no. Electrical contractor Electrical consultants General contractor Building and electrical details Building state: New Existing Proposed use of building: Permanent service Temporary service Number of hours of operation per day: Number of operational days per week: Main switch size: amps Code peak demand: kW Your Designer may inquire further about your computation 80% rated 100% rated (BC Hydro approval required) for the code demand Voltage: volts Average operating demand: kW Largest motor: hp Time of day of peak demand: AM PM Fire pump: No Yes - Size Building heated by: Gas Electric Heat pump - Locked Rotor Amp A Electric furnace Other On-site customer-owned generation: No Yes—open transition Yes—closed transition Requested service type: Overhead Underground Load details Existing (if applicable) New Electric heating load kW kW Lighting load kW kW Heat pump including geothermal kW kW Air conditioning motor load kW kW Electric vehicle charging load kW kW Other motor load kW kW Other load kW kW Total connected load kW kW Proposed future load kW kW Metering Information Total number of meters Residential: Commercial: Voltage: Meter type: 120/240 120/208 347/600 TBD 1 Phase 3 Phase TBD Meter details Other: Temporary master metering required Number of wires: Yes No Current transformer Bar CT lugs conductor size: x Window TBD CT type: Conductor to CT lugs: Cu Al Energization ○ Energization of the project will be scheduled upon receipt of: • Necessary approvals, permits from appropriate authorities, including municipal, electric inspection and other utilities. -

The Story of the Coquitlam River Watershed Past, Present and Future
Fraser Salmon and Watersheds Program – Living Rivers Project Coquitlam River Stakeholder Engagement Phase I The Story of the Coquitlam River Watershed Past, Present and Future Prepared for: The City of Coquitlam and Kwikwetlem First Nation Funding provided by: The Pacific Salmon Foundation Additional funding provided by Fisheries and Oceans Canada Prepared by: Jahlie Houghton, JR Environmental – April 2008 Updated by: Coquitlam River Watershed Work Group – October 2008 Final Report: October 24, 2008 2 File #: 13-6410-01/000/2008-1 Doc #: 692852.v1B Acknowledgements I would like to offer a special thanks to individuals of the community who took the time to meet with me, who not only helped to educate me on historical issues and events in the watershed, but also provided suggestions to their vision of what a successful watershed coordinator could contribute in the future. These people include Elaine Golds, Niall Williams, Don Gillespie, Dianne Ramage, Tony Matahlija, Tim Tyler, John Jakse, Vance Reach, Sherry Carroll, Fin Donnelly, Maurice Coulter-Boisvert, Matt Foy, Derek Bonin, Charlotte Bemister, Dave Hunter, Jim Allard, Tom Vanichuk, and George Turi. I would also like to thank members of the City of Coquitlam, Kwikwetlem First Nation, the Department of Fisheries and Oceans, and Watershed Watch Salmon Society (representative for Kwikwetlem) who made this initiative possible and from whom advice was sought throughout this process. These include Jennifer Wilkie, Dave Palidwor, Mike Carver, Margaret Birch, Hagen Hohndorf, Melony Burton, Tom Cadieux, Dr. Craig Orr, George Chaffee, and Glen Joe. Thank you to the City of Coquitlam also for their printing and computer support services. -

BC Hydro's Response to Intervener Information Requests Round 4
Fred James Chief Regulatory Officer Phone: 604-623-4046 Fax: 604-623-4407 [email protected] November 14, 2019 Mr. Patrick Wruck Commission Secretary and Manager Regulatory Support British Columbia Utilities Commission Suite 410, 900 Howe Street Vancouver, BC V6Z 2N3 Dear Mr. Wruck: RE: Project No. 1598990 British Columbia Utilities Commission (BCUC or Commission) British Columbia Hydro and Power Authority (BC Hydro) Fiscal 2020 to Fiscal 2021 Revenue Requirements Application (the Application) BC Hydro writes in compliance with Commission Order No. G-279-19 to provide its responses to Round 4 information requests as follows: Exhibit B-22 Responses to Commission IRs (Public Version) Exhibit B-22-1 Responses to Commission IRs (Confidential Version) Exhibit B-23 Responses to Intervener IRs (Public Version) Exhibit B-23-1 Responses to Intervener IRs (Confidential Version) BC Hydro is filing a number of IR responses and/or attachments to responses confidentially with the Commission. In each instance, an explanation for the request for confidential treatment is provided in the public version of the IR response. BC Hydro seeks this confidential treatment pursuant to section 42 of the Administrative Tribunals Act and Part 4 of the Commission’s Rules of Practice and Procedure. Overall, BC Hydro received 327 Round 4 information requests. Responses to 260 of those information requests are included in this filing. Thirty-four information requests are the subject of BC Hydro’s letter of November 8, 2019 (the information requests at issue are listed in the attachment to that letter). They are being addressed through the comment process established by Order No. -

Appendix B: Hydrotechnical Assessment
Sheep Paddocks Trail Alignment Analysis APPENDIX B: HYDROTECHNICAL ASSESSMENT LEES+Associates -112- 30 Gostick Place | North Vancouver, BC V7M 3G3 | 604.980.6011 | www.nhcweb.com 300217 15 August 2013 Lees + Associates Landscape Architects #509 – 318 Homer Street Vancouver, BC V6B 2V2 Attention: Nalon Smith Dear Mr. Smith: Subject: Sheep Paddocks Trail Alignment – Phase 1 Hydrotechnical Assessment Preliminary Report 1 INTRODUCTION Metro Vancouver wishes to upgrade the Sheep Paddocks Trail between Pitt River Road and Mundy Creek in Colony Farm Regional Park on the west side of the Coquitlam River. The trail is to accommodate pedestrian and bicycle traffic and be built to withstand at least a 1 in 10 year flood. The project will be completed in three phases: 1. Phase 1 – Route Selection 2. Phase 2 – Detailed Design 3. Phase 3 – Construction and Post-Construction This letter report provides hydrotechnical input for Phase 1 – Route Selection. Currently, a narrow footpath runs along the top of a berm on the right bank of the river. The trail suffered erosion damage in 2007 and was subsequently closed to the public but is still unofficially in use. Potential future routes include both an inland and river option, as well as combinations of the two. To investigate the feasibility of the different options and help identify the most appropriate trail alignment from a hydrotechnical perspective, NHC was retained to undertake the following Phase I scope of work: • Participate in three meetings. • Attend a site visit. • Estimate different return period river flows and comment on local drainage requirements. • Simulate flood levels and velocities corresponding to the different flows. -

BC Hydro and Power Authority 2021/22
BC Hydro and Power Authority 2021/22 – 2023/24 Service Plan April 2021 For more information on BC Hydro contact: 333 Dunsmuir Street Vancouver, BC V6B 5R3 Lower Mainland 604 BCHYDRO (604 224 9376) Outside Lower Mainland 1 800 BCHYDRO (1 800 224 9376)] Or visit our website at bchydro.com Published by BC Hydro BC Hydro and Power Authority Board Chair’s Accountability Statement The 2021/22 – 2023/24 BC Hydro Service Plan was prepared under the Board’s direction in accordance with the Budget Transparency and Accountability Act. The plan is consistent with government’s strategic priorities and fiscal plan. The Board is accountable for the contents of the plan, including what has been included in the plan and how it has been reported. The Board is responsible for the validity and reliability of the information included in the plan. All significant assumptions, policy decisions, events and identified risks, as of February 28, 2021 have been considered in preparing the plan. The performance measures presented are consistent with the Budget Transparency and Accountability Act, BC Hydro’s mandate and goals, and focus on aspects critical to the organization’s performance. The targets in this plan have been determined based on an assessment of BC Hydro’s operating environment, forecast conditions, risk assessment and past performance. Doug Allen Board Chair 2021/22 – 2023/24 Service Plan Page | 3 BC Hydro and Power Authority Table of Contents Board Chair’s Accountability Statement ....................................................................................... -

Bc Hydro Rights of Way Guidelines Compatible Uses and Development Near Power Lines Contents
BC HYDRO RIGHTS OF WAY GUIDELINES COMPATIBLE USES AND DEVELOPMENT NEAR POWER LINES CONTENTS OVERVIEW ..........................................................................................................................................................................................................3 WHO ARE THESE GUIDELINES FOR? ................................................................................................................................................................4 PREPARING AND SUBMITTING A PROPOSAL ..................................................................................................................................................5 POSSIBLE COMPATIBLE USES OF RIGHT OF WAY ...........................................................................................................................................6 GENERAL SAFETY INFORMATION .....................................................................................................................................................................7 PLANTING AND LOGGING NEAR POWER LINES ..............................................................................................................................................8 UNDERGROUND INSTALLATIONS .....................................................................................................................................................................9 DESIGNING AROUND BC HYDRO RIGHTS OF WAY .........................................................................................................................................11 -

Port Coquitlam Flood Mapping Update
Port Coquitlam Flood Mapping Update RECOMMENDATION: None. PREVIOUS COUNCIL/COMMITTEE ACTION On September 17, 2019 Council carried the following motion: That staff prepare flood maps showing current flood risk to Port Coquitlam from the Fraser Basin and provide a report in the fall 2019 with information about the risks facing the community from rising sea levels that align with projections in the most recent Intergovernmental Panel on Climate Change report. REPORT SUMMARY Port Coquitlam has participated in the Fraser Basin Council’s Lower Mainland Flood Management Strategy (“the Strategy”) since its development in 2014. Participants in the strategy have responsibilities or interests related to flood management and include the Government of Canada, the Province of British Columbia, Lower Mainland local governments, First Nations and non- governmental and private sector entities in the region. This report summarizes the flood projections for Port Coquitlam, the regional work completed to date and presents the Strategy’s next phase. BACKGROUND The Lower Fraser Watershed is fed by 12 major watersheds. 1. The Upper / Middle Fraser 7. Chilcotin 2. Stuart 8. North Thompson 3. McGregor 9. South Thompson 4. Nechako 10. Thompson 5. Quesnel 11. Lillooet 6. West Road-Blackwater 12. Harrison These watersheds are illustrated on Figure 1. Report To: Committee of Council Department: Engineering & Public Works Approved by: F. Smith Meeting Date: November 19, 2019 Port Coquitlam Flood Mapping Update Figure 1 – Fraser Basin Watersheds https://www.fraserbasin.bc.ca/basin_watersheds.html In addition, the Lower Fraser watershed incorporates a number of smaller watersheds: Stave Lake and River drain into the Fraser between Maple Ridge and Mission; Alouette Lake and River flow into the Pitt River; the Pitt River drains south from Garibaldi Provincial Park through Pitt Lake, emptying into the Fraser River between Pitt Meadows and Port Coquitlam. -

August 7, 2019 Mr. Patrick Wruck Commission Secretary
Fred James Chief Regulatory Officer Phone: 604-623-4046 Fax: 604-623-4407 [email protected] August 7, 2019 Mr. Patrick Wruck Commission Secretary and Manager Regulatory Support British Columbia Utilities Commission Suite 410, 900 Howe Street Vancouver, BC V6Z 2N3 Dear Mr. Wruck: RE: British Columbia Utilities Commission (BCUC or Commission) British Columbia Hydro and Power Authority (BC Hydro) Fleet Electrification Rate Application BC Hydro writes to file its Application pursuant to sections 58 to 60 of the Utilities Commission Act (UCA) for approval of Rate Schedules 164x – Overnight Rate (150 kW and over) and 165x - Demand Transition Rate (150 kW and over) for use for charging of electric fleet vehicles and vessels. These new services are in response to customer requests for fleet charging rates and support the electrification of commercial fleet vehicles and vessels. For further information, please contact Anthea Jubb at 604-623-3545 or by email at [email protected]. Yours sincerely, (for) Fred James Chief Regulatory Officer ac/tl Enclosure British Columbia Hydro and Power Authority, 333 Dunsmuir Street, Vancouver BC V6B 5R3 www.bchydro.com BC Hydro Fleet Electrification Rate Application August 7, 2019 August 7, 2019 Table of Contents 1 Introduction and Need for Fleet Electrification Rates ......................................... 1 1.1 Purpose of Application .............................................................................. 1 1.2 Need for the Optional Rates ..................................................................... -

Dams and Hydroelectricity in the Columbia
COLUMBIA RIVER BASIN: DAMS AND HYDROELECTRICITY The power of falling water can be converted to hydroelectricity A Powerful River Major mountain ranges and large volumes of river flows into the Pacific—make the Columbia precipitation are the foundation for the Columbia one of the most powerful rivers in North America. River Basin. The large volumes of annual runoff, The entire Columbia River on both sides of combined with changes in elevation—from the the border is one of the most hydroelectrically river’s headwaters at Canal Flats in BC’s Rocky developed river systems in the world, with more Mountain Trench, to Astoria, Oregon, where the than 470 dams on the main stem and tributaries. Two Countries: One River Changing Water Levels Most dams on the Columbia River system were built between Deciding how to release and store water in the Canadian the 1940s and 1980s. They are part of a coordinated water Columbia River system is a complex process. Decision-makers management system guided by the 1964 Columbia River Treaty must balance obligations under the CRT (flood control and (CRT) between Canada and the United States. The CRT: power generation) with regional and provincial concerns such as ecosystems, recreation and cultural values. 1. coordinates flood control 2. optimizes hydroelectricity generation on both sides of the STORING AND RELEASING WATER border. The ability to store water in reservoirs behind dams means water can be released when it’s needed for fisheries, flood control, hydroelectricity, irrigation, recreation and transportation. Managing the River Releasing water to meet these needs influences water levels throughout the year and explains why water levels The Columbia River system includes creeks, glaciers, lakes, change frequently. -

Columbia Basin Plan
FOR REFERENCE ONLY This version is now archived. Updated 2019 Columbia Region Action Plans available at: fwcp.ca/region/columbia-region Photo credit: Larry Halverson COLUMBIA BASIN PLAN June 2012 Contents 1. Introduction ......................................................................................................................... 1 1.1 Fish and Wildlife Compensation Program ........................................................................ 1 Vision ........................................................................................................................................ 2 Principles .................................................................................................................................. 2 Partners .................................................................................................................................... 2 Policy Context ........................................................................................................................... 2 Program Delivery ...................................................................................................................... 4 Project Investment Criteria ...................................................................................................... 4 2. The Columbia River Basin .................................................................................................... 6 2.1 Setting ............................................................................................................................. -

Building of the Coquitlam River and Port Moody Trails Researched and Written by Ralph Drew, Belcarra, BC, June 2010; Updated Dec 2012 and Dec 2013
Early Trail Building in the New Colony of British Columbia — John Hall’s Building of the Coquitlam River and Port Moody Trails Researched and written by Ralph Drew, Belcarra, BC, June 2010; updated Dec 2012 and Dec 2013. A recent “find” of colonial correspondence in the British Columbia Archives tells a story about the construction of the Coquitlam River and Port Moody Trails between 1862 and 1864 by pioneer settler John Hall. (In 1870 Hall pre-empted 160 acres of Crown Land on Indian Arm and became Belcarra’s first European settler.) The correspondence involves a veritable “who’s who” of people in the administration in the young ‘Colony of British Columbia’. This historic account serves to highlight one of the many challenges faced by our pioneers during the period of colonial settlement in British Columbia. Sir James Douglas When the Fraser River Gold Rush began in the spring of 1858, there were only about 250 to 300 Europeans living in the Fraser Valley. The gold rush brought on the order of 30,000 miners flocking to the area in the quest for riches, many of whom came north from the California gold fields. As a result, the British Colonial office declared a new Crown colony on the mainland called ‘British Columbia’ and appointed Sir James Douglas as the first Governor. (1) The colony was first proclaimed at Fort Langley on 19th November, 1858, but in early 1859 the capital was moved to the planned settlement called ‘New Westminster’, Sir James Douglas strategically located on the northern banks of the Fraser River. -

BC Hydro Climate Change Assessment Report 2012
POTENTIAL IMPACTS OF CLIMATE CHANGE ON BC HYDRO’S WATER RESOURCES Georg Jost: Ph.D., Senior Hydrologic Modeller, BC Hydro Frank Weber; M.Sc., P. Geo., Lead, Runoff Forecasting, BC Hydro 1 EXecutiVE Summary Global climate change is upon us. Both natural cycles and anthropogenic greenhouse gas emissions influence climate in British Columbia and the river flows that supply the vast majority of power that BC Hydro generates. BC Hydro’s climate action strategy addresses both the mitigation of climate change through reducing our greenhouse gas emissions, and adaptation to climate change by understanding the risks and magnitude of potential climatic changes to our business today and in the future. As part of its climate change adaptation strategy, BC Hydro has undertaken internal studies and worked with some of the world’s leading scientists in climatology, glaciology, and hydrology to determine how climate change affects water supply and the seasonal timing of reservoir inflows, and what we can expect in the future. While many questions remain unanswered, some trends are evident, which we will explore in this document. 2 IMPACTS OF CLIMATE CHANGE ON BC HYDRO-MANAGED WATER RESOURCES W HAT we haVE seen so far » Over the last century, all regions of British Columbia »F all and winter inflows have shown an increase in became warmer by an average of about 1.2°C. almost all regions, and there is weaker evidence »A nnual precipitation in British Columbia increased by for a modest decline in late-summer flows for those about 20 per cent over the last century (across Canada basins driven primarily by melt of glacial ice and/or the increases ranged from 5 to 35 per cent).