Phyton Annales Rei Botanicae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Allium Macleanii Baker, 1883
Allium macleanii Baker, 1883 Identifiants : 1573/allmac Association du Potager de mes/nos Rêves (https://lepotager-demesreves.fr) Fiche réalisée par Patrick Le Ménahèze Dernière modification le 30/09/2021 Classification phylogénétique : Clade : Angiospermes ; Clade : Monocotylédones ; Ordre : Asparagales ; Famille : Amaryllidaceae ; Classification/taxinomie traditionnelle : Règne : Plantae ; Sous-règne : Tracheobionta ; Division : Magnoliophyta ; Classe : Liliopsida ; Ordre : Liliales ; Famille : Amaryllidaceae ; Sous-famille : Allioideae ; Tribu : Allieae ; Genre : Allium ; Synonymes : x (=) basionym, Allium elatum Regel 1884 ; Nom(s) anglais, local(aux) et/ou international(aux) : Oriental royal Salep, royal salep , kabullök (sv) ; Note comestibilité : *** Rapport de consommation et comestibilité/consommabilité inférée (partie(s) utilisable(s) et usage(s) alimentaire(s) correspondant(s)) : Racine (bulbes : crus ou cuits [nourriture(dp*)µ/alimentµ27(+x) et/ou assaisonnement : aromate (fines-herbes et/ou{{{(dp*) condiment27(+x) aromatique{{{(dp*))]) et feuille (feuilles : idem bulbes|{{{(dp*)(27(+x)), ex. comme potherbe{{{(dp*)) comestibles.(1*) Les bulbes sont consommés crus ou cuits. Les feuilles sont consommées crues ou cuites. Les fleurs sont utilisées crues pour parfumer les salades Partie testée : bulbe{{{0(+x) (traduction automatique) Original : Bulb{{{0(+x) Taux d'humidité Énergie (kj) Énergie (kcal) Protéines (g) Pro- Vitamines C (mg) Fer (mg) Zinc (mg) vitamines A (µg) 0 0 0 0 0 0 0 (1*)Voir genre Allium pour les précautions à prendre (risques de confusion et possible toxicité à fortes doses).(1*)Voir genre Allium pour les précautions à prendre (risques de confusion et possible toxicité à fortes doses){{{(rp*). Note médicinale : ** Illustration(s) (photographie(s) et/ou dessin(s)): Page 1/3 Autres infos : Plante importante localement{{{27(+x). -

– the 2020 Horticulture Guide –
– THE 2020 HORTICULTURE GUIDE – THE 2020 BULB & PLANT MART IS BEING HELD ONLINE ONLY AT WWW.GCHOUSTON.ORG THE DEADLINE FOR ORDERING YOUR FAVORITE BULBS AND SELECTED PLANTS IS OCTOBER 5, 2020 PICK UP YOUR ORDER OCTOBER 16-17 AT SILVER STREET STUDIOS AT SAWYER YARDS, 2000 EDWARDS STREET FRIDAY, OCTOBER 16, 2020 SATURDAY, OCTOBER 17, 2020 9:00am - 5:00pm 9:00am - 2:00pm The 2020 Horticulture Guide was generously underwritten by DEAR FELLOW GARDENERS, I am excited to welcome you to The Garden Club of Houston’s 78th Annual Bulb and Plant Mart. Although this year has thrown many obstacles our way, we feel that the “show must go on.” In response to the COVID-19 situation, this year will look a little different. For the safety of our members and our customers, this year will be an online pre-order only sale. Our mission stays the same: to support our community’s green spaces, and to educate our community in the areas of gardening, horticulture, conservation, and related topics. GCH members serve as volunteers, and our profits from the Bulb Mart are given back to WELCOME the community in support of our mission. In the last fifteen years, we have given back over $3.5 million in grants to the community! The Garden Club of Houston’s first Plant Sale was held in 1942, on the steps of The Museum of Fine Arts, Houston, with plants dug from members’ gardens. Plants propagated from our own members’ yards will be available again this year as well as plants and bulbs sourced from near and far that are unique, interesting, and well suited for area gardens. -

(Curculigo Orchioides) and Salam (Eulophia Compestris) Madan B
Int J Ayu Pharm Chem REVIEW ARTICLE www.ijapc.com e-ISSN 2350-0204 A Review Article on Species used as Musali (Curculigo orchioides) and Salam (Eulophia compestris) Madan B. Tonge* *Dept. of Dravyaguna, Govt. Ayurved College Nanded (MS), India Abstract In day to day practice when we see the market samples of Musali it creates confusion in mind; which type Musali is sold by the vendor. These days various species of plants are used as Musali in different parts of India. Traditionally, Salam and Salam panja are also used as Mushali. To rule out all these differences and arrive to a definite conclusion. This is an attempt to collect the referances from samhitas and nighantus about musali. Botanically classify the species which are used as musali. Describe all the species which are in use as musali in a systematic manner. Keywords Mushali, Shweta Musali, Salam, Talmuli Greentree Group Received 09/08/16 Accepted 29/08/16 Published 10/09/16 ________________________________________________________________________________________________________ Madan B Tonge 2016 Greentree Group © IJAPC Int J Ayu Pharm Chem 2016 Vol. 5 Issue 2 www.ijapc.com 283 [e ISSN 2350-0204] Int J Ayu Pharm Chem INTRODUCTION shwveta musali few species of asparagaceae The term musali is famous in traditional family are in use and also roots of salam Indian system of medicine. Medicine with mishri and salampanja mishri are used as musali name is known to many household in musali. The word mishri is derived from India. Most commonly used as a tonic, musali, so few people call it as salam aphrodisiac, rejuvenator for increasing musali, salam panja musali. -

Hill View Rare Plants, Summer Catalogue 2011, Australia
Summer 2011/12 Hill View Rare Plants Calochortus luteus Calochortus superbus Susan Jarick Calochortus albidus var. rubellus 400 Huon Road South Hobart Tas 7004 Ph 03 6224 0770 Summer 2011/12 400 Huon Road South Hobart Tasmania, 7004 400 Huon Road South Hobart Tasmania, 7004 Summer 2011/12 Hill View Rare Plants Ph 03 6224 0770 Ph 03 6224 0770 Hill View Rare Plants Marcus Harvey’s Hill View Rare Plants 400 Huon Road South Hobart Tasmania, 7004 Welcome to our 2011/2012 summer catalogue. We have never had so many problems in fitting the range of plants we have “on our books” into the available space! We always try and keep our lists “democratic” and balanced although at times our prejudices show and one or two groups rise to the top. This year we are offering an unprecedented range of calochortus in a multiplicity of sizes, colours and flower shapes from the charming fairy lanterns of C. albidus through to the spectacular, later-flowering mariposas with upward-facing bowl-shaped flowers in a rich tapestry of shades from canary-yellow through to lilac, lavender and purple. Counterpoised to these flashy dandies we are offering an assortment of choice muscari whose quiet charm, softer colours and Tulipa vvedenskyi Tecophilaea cyanocrocus Violacea persistent flowering make them no less effective in the winter and spring garden. Standouts among this group are the deliciously scented duo, M. muscarimi and M. macrocarpum and the striking and little known tassel-hyacith, M. weissii. While it has its devotees, many gardeners are unaware of the qualities of the large and diverse tribe of “onions”, known as alliums. -

Van Zyverden's
Van Zyverden’s ALLIUM KARATAVIENSE Allium are in the same family as garlic, onions, chives and shallots. This makes gardeners wonder if they should include them in their ornamental gardening plans, as it conjures up images of supermarket produce. But because good garden designs are often made up of different shapes, allium’s rounded blooms make for high drama and interest in the garden. The Allium group gets more popular annually, from over 300 species to choose. They amaze everyone, and few plants create this kind of wow in the garden. We will be adding many new varieties shortly. Leaves and bulbs Commonly called Turkistan onion Deer and rodent resistant have a mild onion-like aroma when cut or bruised. About This Variety: Allium Karataviense is a compact, bulbous perennial that is ornamentally grown for both its foliage and its flowers. It is native to the Karatau Mountains (hence the specific epithet) in Kazakhstan. Broad-elliptic, spreading, gray-green, basal leaves appear in pairs. Leaves are sometimes mottled with purple. In late spring, a short but sturdy flowering stem rises from the center of each leaf pair. Each flowering stem is topped with a large spherical flower head containing tiny, star-shaped, dull pink florets. Flowers bloom in early summer. Flowers have a mild fragrance. Growing Instructions: As Alliums do not like wet feet, find a sunny location where the soil drains well or try to improve the drainage. The bulbs will rot in wet areas. Aside from that, almost no maintenance is required. Care Tip: Dig, divide, and replant bulbs after a few years of decreasing flower production. -

Aksu-Zhabagly BIOSPHERE RESERVE National Commission Republic of Kazakhstan
Aksu-Zhabagly BIOSPHERE RESERVE National Commission Republic of Kazakhstan Kazakhstan National Committee Kazakhstan National Committee for the UNESCO Programme “Man and Biosphere” MAB, Institute of Zoology, 93 al-Farabi Str. Almaty, 050060 KAZAKHSTAN Kazakhstan National Committee Aksu-Zhabagly Biosphere Reserve NominatioN PART I: SUMMARY 1. PROPOSED NAME OF THE BIOSPHERE RESERVE: Aksu-Zhabagly Biosphere Reserve 2. COUNTRY: Kazakhstan Aksu-Zhabagly 4 FULFILLMENT OF THE THREE FUNCTIONS OF BIOSPHERE RESERVES 3. «Conservation — contribute to the conservation of landscapes, ecosystems, species and genetic variation» 3. 1 Aksu-Zhabagly biosphere reserve is located in the Western end of Talasskiy Alatau and Southern part of Karatau in the West Tien Shan. The whole region of the West Tien Shan is an Eastern outpost of Mediterranean atmospheric circulation, therefore it has a winter-spring rainfall. The mountain range of the West Tien Shan is a barrier that catches the moisture in the Western transport of air masses; in addition, this region is situated within the zone of the Southern deserts, where the annual temperature sum is high and about 4000-5000o C. As a result, this area is the most favorable for vegetation and preservation of many ancient relict species and plant communities. Moreover, the reserve’s ecosystems have a very close relationship with the natural systems of the Near East and the Mediterranean than to the rest of the ecosystems of the Tien Shan. The territory of Aksu Zhabagly has a high degree of representativeness at regional level. For example, it has almost all landscape types and sub-types of the West Tien Shan, except for deserts and gypsophilous subshrub communities, which are well below the reserve in altitude. -

ISTA List of Stabilized Plant Names 7Th Edition
ISTA List of Stabilized Plant Names th 7 Edition ISTA Nomenclature Committee Chair: Dr. M. Schori Published by All rights reserved. No part of this publication may be The Internation Seed Testing Association (ISTA) reproduced, stored in any retrieval system or transmitted Zürichstr. 50, CH-8303 Bassersdorf, Switzerland in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without prior ©2020 International Seed Testing Association (ISTA) permission in writing from ISTA. ISBN 978-3-906549-77-4 ISTA List of Stabilized Plant Names 1st Edition 1966 ISTA Nomenclature Committee Chair: Prof P. A. Linehan 2nd Edition 1983 ISTA Nomenclature Committee Chair: Dr. H. Pirson 3rd Edition 1988 ISTA Nomenclature Committee Chair: Dr. W. A. Brandenburg 4th Edition 2001 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 5th Edition 2007 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 6th Edition 2013 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 7th Edition 2019 ISTA Nomenclature Committee Chair: Dr. M. Schori 2 7th Edition ISTA List of Stabilized Plant Names Content Preface .......................................................................................................................................................... 4 Acknowledgements ....................................................................................................................................... 6 Symbols and Abbreviations .......................................................................................................................... -

Classical and Hebrew Sages on Cultivated Biennial Plants Part II
Classical and Hebrew Sages on Cultivated Biennial Plants Part II Moshe Negbi, Eliezer Ε. Goldschmidt and Nikolaj Serikoff Liliaceae Allium Theophrastus, with his profound botanical understanding, grouped together the members of the genus Allium known to him: onion (kromnon, A. cepa), leek (prason, A. porrum), garlic (skordon, A. sativum), long onion (gethnon, A. cepa var.1), horn onion (geteion, A. cepa var.) and shallot (kromnon to skhiston, A. ascalonicum). Moreover, in morphologi cal and developmental aspects he compared the Allium species to other monocotyledon- ous geophytes.2 Following Theophrastus we deal with them together. On seed, sowing, germination and vegetative reproduction I: Seed does not keep well ... among the more vigorous ones are ... leek ... among the less vigorous are long onion — which will not keep ... (HP 7.5.5). The second period [of sowing] begins after the winter solstice in the month of Gamelion (January), in which they scatter or plant the seed of leeks, celery, long onion and orach ... Leek and long onion do not take the same time [to germinate], but the former 19 to 20 days, the latter 10 to 12 (HP 7Ἰ.2-3). Another thing that makes a difference as to the ra pidity with which the seeds germinate is their age; for some herbs come up quicker from fresh seed, as leek, long onion ... (HP 7.1.6). All these herbs are propagated from seed, and some also by ... a piece of root... By root [bulb!] are planted garlic, onion ... and in general such bulbous plants (HP 7.2Ἰ).3 On their leaves: ... in most potherbs [leaves] grow directly from the root, as in onion, garlic, chicory, and also in asphodel, squill and purse-tassels .. -

Listado De Todas Las Plantas Que Tengo Fotografiadas Ordenado Por Familias Según El Sistema APG III (Última Actualización: 2 De Septiembre De 2021)
Listado de todas las plantas que tengo fotografiadas ordenado por familias según el sistema APG III (última actualización: 2 de Septiembre de 2021) GÉNERO Y ESPECIE FAMILIA SUBFAMILIA GÉNERO Y ESPECIE FAMILIA SUBFAMILIA Acanthus hungaricus Acanthaceae Acanthoideae Metarungia longistrobus Acanthaceae Acanthoideae Acanthus mollis Acanthaceae Acanthoideae Odontonema callistachyum Acanthaceae Acanthoideae Acanthus spinosus Acanthaceae Acanthoideae Odontonema cuspidatum Acanthaceae Acanthoideae Aphelandra flava Acanthaceae Acanthoideae Odontonema tubaeforme Acanthaceae Acanthoideae Aphelandra sinclairiana Acanthaceae Acanthoideae Pachystachys lutea Acanthaceae Acanthoideae Aphelandra squarrosa Acanthaceae Acanthoideae Pachystachys spicata Acanthaceae Acanthoideae Asystasia gangetica Acanthaceae Acanthoideae Peristrophe speciosa Acanthaceae Acanthoideae Barleria cristata Acanthaceae Acanthoideae Phaulopsis pulchella Acanthaceae Acanthoideae Barleria obtusa Acanthaceae Acanthoideae Pseuderanthemum carruthersii ‘Rubrum’ Acanthaceae Acanthoideae Barleria repens Acanthaceae Acanthoideae Pseuderanthemum carruthersii var. atropurpureum Acanthaceae Acanthoideae Brillantaisia lamium Acanthaceae Acanthoideae Pseuderanthemum carruthersii var. reticulatum Acanthaceae Acanthoideae Brillantaisia owariensis Acanthaceae Acanthoideae Pseuderanthemum laxiflorum Acanthaceae Acanthoideae Brillantaisia ulugurica Acanthaceae Acanthoideae Pseuderanthemum laxiflorum ‘Purple Dazzler’ Acanthaceae Acanthoideae Crossandra infundibuliformis Acanthaceae Acanthoideae Ruellia -
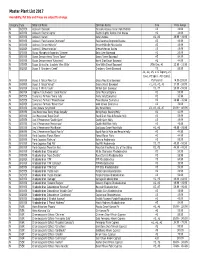
Master Plant List 2017.Xlsx
Master Plant List 2017 Availability, Pot Size and Prices are subject to change. Category Type Botanical Name Common Name Size Price Range N BREVER Azalea X 'Cascade' Cascade Azalea (Glenn Dale Hybrid) #3 49.99 N BREVER Azalea X 'Electric Lights' Electric Lights Double Pink Azalea #2 44.99 N BREVER Azalea X 'Karen' Karen Azalea #2, #3 39.99 - 49.99 N BREVER Azalea X 'Poukhanense Improved' Poukhanense Improved Azalea #3 49.99 N BREVER Azalea X 'Renee Michelle' Renee Michelle Pink Azalea #3 49.99 N BREVER Azalea X 'Stewartstonian' Stewartstonian Azalea #3 49.99 N BREVER Buxus Microphylla Japonica "Gregem' Baby Gem Boxwood #2 29.99 N BREVER Buxus Sempervirens 'Green Tower' Green Tower Boxwood #5 64.99 N BREVER Buxus Sempervirens 'Katerberg' North Star Dwarf Boxwood #2 44.99 N BREVER Buxus Sinica Var. Insularis 'Wee Willie' Wee Willie Dwarf Boxwood Little One, #1 13.99 - 21.99 N BREVER Buxus X 'Cranberry Creek' Cranberry Creek Boxwood #3 89.99 #1, #2, #5, #15 Topiary, #5 Cone, #5 Spiral, #10 Spiral, N BREVER Buxus X 'Green Mountain' Green Mountain Boxwood #5 Pyramid 14.99-299.99 N BREVER Buxus X 'Green Velvet' Green Velvet Boxwood #1, #2, #3, #5 17.99 - 59.99 N BREVER Buxus X 'Winter Gem' Winter Gem Boxwood #5, #7 59.99 - 99.99 N BREVER Daphne X Burkwoodii 'Carol Mackie' Carol Mackie Daphne #2 59.99 N BREVER Euonymus Fortunei 'Ivory Jade' Ivory Jade Euonymus #2 35.99 N BREVER Euonymus Fortunei 'Moonshadow' Moonshadow Euonymus #2 29.99 - 35.99 N BREVER Euonymus Fortunei 'Rosemrtwo' Gold Splash Euonymus #2 39.99 N BREVER Ilex Crenata 'Sky Pencil' -

Survey of Wild Food Plants for Human Consumption in Geçitli (Hakkari, Turkey)
Indian Journal of Traditional Knowledge Vol. 14(2), April 2015, pp. 183-190 Survey of wild food plants for human consumption in Geçitli (Hakkari, Turkey) İdris Kaval1, Lütfi Behçet2 & Uğur Çakilcioğlu3* 1Yuzuncu Yıl University, Department of Biology, Van 65000, Turkey; 2Bingöl University, Department of Biology, Bingöl 12000, Turkey; 3Tunceli University, Pertek Sakine Genç Vocational School, Pertek, Tunceli 62500, Turkey E-mails: [email protected]; [email protected]; [email protected] Received 15 July 2014, revised 22 January 2015 This study aims to record accumulation of knowledge on plants which are used as food by native people of Geçitli (Hakkari, Turkey) that has a rich culture and a very natural environment. In addition, the medical uses of these plants were compiled from the literature. Study area was located on the East of Anatolian diagonal, in the Eastern Anatolia region. Field study was carried out over a period of approximately two years (2008-2010). During this period, 84 vascular plant taxa were collected. The plants were pressed in the field and prepared for identification. A total of 84 food plants belonging to 30 families were identified in the region. In the study being conducted, use of wild plants as food points out interest of people in Geçitli in wild plants. The fact that a large proportion of edible plants are also being used for medicinal purposes indicates that the use of wild plants has a high potential in the region. The present study shows that further ethnobotanical investigations are worthy to be carried out in Turkey, where most of knowledge on popular food plants are still to discover. -

Table of Contents
WELCOME TO LOST HORIZONS 2015 CATALOGUE Table of Contents Welcome to Lost Horizons . .15 . Great Plants/Wonderful People . 16. Nomenclatural Notes . 16. Some History . 17. Availability . .18 . Recycle . 18 Location . 18 Hours . 19 Note on Hardiness . 19. Gift Certificates . 19. Lost Horizons Garden Design, Consultation, and Construction . 20. Understanding the catalogue . 20. References . 21. Catalogue . 23. Perennials . .23 . Acanthus . .23 . Achillea . .23 . Aconitum . 23. Actaea . .24 . Agastache . .25 . Artemisia . 25. Agastache . .25 . Ajuga . 26. Alchemilla . 26. Allium . .26 . Alstroemeria . .27 . Amsonia . 27. Androsace . .28 . Anemone . .28 . Anemonella . .29 . Anemonopsis . 30. Angelica . 30. For more info go to www.losthorizons.ca - Page 1 Anthericum . .30 . Aquilegia . 31. Arabis . .31 . Aralia . 31. Arenaria . 32. Arisaema . .32 . Arisarum . .33 . Armeria . .33 . Armoracia . .34 . Artemisia . 34. Arum . .34 . Aruncus . .35 . Asarum . .35 . Asclepias . .35 . Asparagus . .36 . Asphodeline . 36. Asphodelus . .36 . Aster . .37 . Astilbe . .37 . Astilboides . 38. Astragalus . .38 . Astrantia . .38 . Aubrieta . 39. Aurinia . 39. Baptisia . .40 . Beesia . .40 . Begonia . .41 . Bergenia . 41. Bletilla . 41. Boehmeria . .42 . Bolax . .42 . Brunnera . .42 . For more info go to www.losthorizons.ca - Page 2 Buphthalmum . .43 . Cacalia . 43. Caltha . 44. Campanula . 44. Cardamine . .45 . Cardiocrinum . 45. Caryopteris . .46 . Cassia . 46. Centaurea . 46. Cephalaria . .47 . Chelone . .47 . Chelonopsis . ..