Ventilatory Action of the Hypaxial Muscles of the Lizard Iguana Iguana: a Function of Slow Muscle
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
The Structure and Function of Breathing
CHAPTERCONTENTS The structure-function continuum 1 Multiple Influences: biomechanical, biochemical and psychological 1 The structure and Homeostasis and heterostasis 2 OBJECTIVE AND METHODS 4 function of breathing NORMAL BREATHING 5 Respiratory benefits 5 Leon Chaitow The upper airway 5 Dinah Bradley Thenose 5 The oropharynx 13 The larynx 13 Pathological states affecting the airways 13 Normal posture and other structural THE STRUCTURE-FUNCTION considerations 14 Further structural considerations 15 CONTINUUM Kapandji's model 16 Nowhere in the body is the axiom of structure Structural features of breathing 16 governing function more apparent than in its Lung volumes and capacities 19 relation to respiration. This is also a region in Fascla and resplrstory function 20 which prolonged modifications of function - Thoracic spine and ribs 21 Discs 22 such as the inappropriate breathing pattern dis- Structural features of the ribs 22 played during hyperventilation - inevitably intercostal musculature 23 induce structural changes, for example involving Structural features of the sternum 23 Posterior thorax 23 accessory breathing muscles as well as the tho- Palpation landmarks 23 racic articulations. Ultimately, the self-perpetuat- NEURAL REGULATION OF BREATHING 24 ing cycle of functional change creating structural Chemical control of breathing 25 modification leading to reinforced dysfunctional Voluntary control of breathing 25 tendencies can become complete, from The autonomic nervous system 26 whichever direction dysfunction arrives, for Sympathetic division 27 Parasympathetic division 27 example: structural adaptations can prevent NANC system 28 normal breathing function, and abnormal breath- THE MUSCLES OF RESPIRATION 30 ing function ensures continued structural adap- Additional soft tissue influences and tational stresses leading to decompensation. -

Ch22: Actions of the Respiratory Muscles
CHAPTER 22 ACTIONS OF THE RESPIRATORY MUSCLES André de Troyer he so-called respiratory muscles are those muscles that caudally and ventrally from their costotransverse articula- Tprovide the motive power for the act of breathing. Thus, tions, such that their ventral ends and the costal cartilages although many of these muscles are involved in a variety of are more caudal than their dorsal parts (Figure 22-2B, C). activities, such as speech production, cough, vomiting, and When the ribs are displaced in the cranial direction, their trunk motion, their primary task is to displace the chest wall ventral ends move laterally and ventrally as well as cra- rhythmically to pump gas in and out of the lungs. The pres- nially, the cartilages rotate cranially around the chon- ent chapter, therefore, starts with a discussion of the basic drosternal junctions, and the sternum is displaced ventrally. mechanical structure of the chest wall in humans. Then, the Consequently, there is usually an increase in both the lateral action of each group of muscles is analyzed. For the sake of and the dorsoventral diameters of the rib cage (see Figure clarity, the functions of the diaphragm, the intercostal mus- 22-2B, C). Conversely, a displacement of the ribs in the cau- cles, the muscles of the neck, and the muscles of the abdom- dal direction is usually associated with a decrease in rib cage inal wall are analyzed sequentially. However, since all these diameters. As a corollary, the muscles that elevate the ribs as muscles normally work together in a coordinated manner, their primary action have an inspiratory effect on the rib the most critical aspects of their mechanical interactions are cage, whereas the muscles that lower the ribs have an expi- also emphasized. -

Respiratory Function of the Rib Cage Muscles
Copyright @ERS Journals Ltd 1993 Eur Respir J, 1993, 6, 722-728 European Respiratory Journal Printed In UK • all rights reserved ISSN 0903 • 1936 REVIEW Respiratory function of the rib cage muscles J.N. Han, G. Gayan-Ramirez, A. Dekhuijzen, M. Decramer Respiratory function of the rib cage muscles. J.N. Han, G. Gayan-Ramirez, R. Respiratory Muscle Research Unit, Labo Dekhuijzen, M. Decramer. ©ERS Journals Ltd 1993. ratory of Pneumology, Respiratory ABSTRACT: Elevation of the ribs and expansion of the rib cage result from the Division, Katholieke Universiteit Leuven, co-ordinated action of the rib cage muscles. We wished to review the action and Belgium. interaction of the rib cage muscles during ventilation. Correspondence: M. Decramer The parasternal intercostal muscles appear to play a predominant role during Respiratory Division quiet breathing, both in humans and in anaesthetized dogs. In humans, the para University Hospital sternal intercostals act in concert with the scalene muscles to expand the upper rib Weligerveld 1 cage, and/or to prevent it from being drawn inward by the action of the diaphragm. B-3212 Pellenberg The external intercostal muscles are considered to be active mainly during inspira Leuven tion, and the internal intercostal muscles during expiration. Belgium The respiratory activity of the external intercostals is minimal during quiet breathing both in man and in dogs, but increases with increasing ventilation. In Keywords: Chest wall mechanics contractile properties spiratory activity in the external intercostals can be enhanced in anaesthetized ani rib cage muscles mals and humans by inspiratory mechanical loading and by col stimulation, rib displacement suggesting that the external intercostals may constitute a reserve system, that may be recruited when the desired expansion of the rib cage is increased. -

Pectoral Region and Axilla Doctors Notes Notes/Extra Explanation Editing File Objectives
Color Code Important Pectoral Region and Axilla Doctors Notes Notes/Extra explanation Editing File Objectives By the end of the lecture the students should be able to : Identify and describe the muscles of the pectoral region. I. Pectoralis major. II. Pectoralis minor. III. Subclavius. IV. Serratus anterior. Describe and demonstrate the boundaries and contents of the axilla. Describe the formation of the brachial plexus and its branches. The movements of the upper limb Note: differentiate between the different regions Flexion & extension of Flexion & extension of Flexion & extension of wrist = hand elbow = forearm shoulder = arm = humerus I. Pectoralis Major Origin 2 heads Clavicular head: From Medial ½ of the front of the clavicle. Sternocostal head: From; Sternum. Upper 6 costal cartilages. Aponeurosis of the external oblique muscle. Insertion Lateral lip of bicipital groove (humerus)* Costal cartilage (hyaline Nerve Supply Medial & lateral pectoral nerves. cartilage that connects the ribs to the sternum) Action Adduction and medial rotation of the arm. Recall what we took in foundation: Only the clavicular head helps in flexion of arm Muscles are attached to bones / (shoulder). ligaments / cartilage by 1) tendons * 3 muscles are attached at the bicipital groove: 2) aponeurosis Latissimus dorsi, pectoral major, teres major 3) raphe Extra Extra picture for understanding II. Pectoralis Minor Origin From 3rd ,4th, & 5th ribs close to their costal cartilages. Insertion Coracoid process (scapula)* 3 Nerve Supply Medial pectoral nerve. 4 Action 1. Depression of the shoulder. 5 2. Draw the ribs upward and outwards during deep inspiration. *Don’t confuse the coracoid process on the scapula with the coronoid process on the ulna Extra III. -

On the Anatomy of Intercostal Spaces in Man and Certain Other Mammals1 by Prof
ON THE ANATOMY OF INTERCOSTAL SPACES IN MAN AND CERTAIN OTHER MAMMALS1 BY PROF. M. A. H. SIDDIQI, M.B., D.L.O., M.S., F.R.C.S. (ENG.) AND DR A. N. MULLICK, M.B., B.S. Anatomy Department, King George's Medical College, Lucknow (India) TIHE standard description of the anatomy of the intercostal space has been discussed by Stibbe in a paper recently published in this Journal(2,3). Prof. Walmsley in 1916(1) showed that the intercostal nerves do not lie in the plane between the internal and external intercostal muscles but deep to the internal intercostal, and that they are separated from the pleura by a deeper musculo-fascial plane consisting of subcostal, intercostal and transversus thoracis muscles from behind forwards. According to Davies, Gladstone and Stibbe (3) there are four musculo-fascial planes in each space and in each space the main nerve lies with a collateral nerve deep to the internal intercostal. As the above paper effected a change in the teaching of the anatomy of intercostal space, we carried out the following investigations on human as well as on certain other Mammalian intercostal spaces. DISSECTION OF HUMAN INTERCOSTAL SPACES Sixty thoraces of different ages were dissected. From some of them the intercostal spaces were cut out en bloc to facilitate dissection; in others the thoracic wall was dissected as a whole. In the case of the foetuses microscopic sections were made to locate the muscular planes and the nerves. The results of our dissection were as follows: I. Intercostal muscles (fig. -

Postural and Ventilatory Functions of Intercostal Muscles
ACTA NEUROBIOL. EXP. 1973, 33: 355-380 Lecture delivered at Symposium "Neural control of breathing" held in Warszawa, August 1971 POSTURAL AND VENTILATORY FUNCTIONS OF INTERCOSTAL MUSCLES B. DURON Laboratory of Neurophysiology, Faculty of Medicine, Amiens, France Abstract. During spontaneous breathing, the interchondral muscles present a pat- tern of activity similar to that of the diaphragm. The external intercostals and most of the internal intercostals generally show electrical discharges not related to ventilatory rhythm. Studies of the electrical responses of these muscles in experi- mental variations of their length show that the external and internal intercostak are readily activated by this category of reflexes while the diaphragm and the interchondrals are not. Bilateral multisegmental sections of spinal dorsal roots do not affect the respiratory activity of the diaphragm and of the interchondral muscles; on the contrary, all types of activity - spontaneous or reflex - disappear from the intercostals. Electrical stimulation of appropriate points in the bulbar pyramids in decerebrate cats can activate at the same time different intercostals and leg muscles without modifying the rhythmic inspiratory activity of the diaphragm and the interchondrals. In preparations with chronically implanted electrodes, the inter- costal muscles are chiefly involved in posture. These results fit very well with our histological findings which disclose a much greater density of muscle spindles in external intercostals than in the diaphragm or in the interchondral muscles. INTRODUCTION Our conception of the role played by the intercostal muscles has been dominated for a long time by the anatomical concept of the "thoracic cage" starting with the original mechanical explanation of Hamberger (1748) based on geometric considerations. -

The Anatomy and Function of the Equine Thoracolumbar Longissimus Dorsi Muscle
Aus dem Veterinärwissenschaftlichen Department der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Lehrstuhl für Anatomie, Histologie und Embryologie Vorstand: Prof. Dr. Dr. Fred Sinowatz Arbeit angefertigt unter der Leitung von Dr. Renate Weller, PhD, MRCVS The Anatomy and Function of the equine thoracolumbar Longissimus dorsi muscle Inaugural-Dissertation zur Erlangung der tiermedizinischen Doktorwürde der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Vorgelegt von Christina Carla Annette von Scheven aus Düsseldorf München 2010 2 Gedruckt mit der Genehmigung der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München Dekan: Univ.-Prof. Dr. Joachim Braun Berichterstatter: Priv.-Doz. Dr. Johann Maierl Korreferentin: Priv.-Doz. Dr. Bettina Wollanke Tag der Promotion: 24. Juli 2010 3 Für meine Familie 4 Table of Contents I. Introduction................................................................................................................ 8 II. Literature review...................................................................................................... 10 II.1 Macroscopic anatomy ............................................................................................. 10 II.1.1 Comparative evolution of the body axis ............................................................ 10 II.1.2 Axis of the equine body ..................................................................................... 12 II.1.2.1 Vertebral column of the horse.................................................................... -

The Expiratory Muscles in Tetraplegia
Paraplegia 29 (1991) 359-363 © 1991 International Medical Society of Paraplegia Paraplegia J Review Article: The Expiratory Muscles in Tetraplegia A. De Troyer, MD, M. Estenne, MD Respiratory Research Unit and Chest Service, Erasme University Hospital, Brussels School of Medicine, Brussels, Belgium. All the well-recognised muscles of expiration, such as the muscles of the anterolate ral wall of the abdomen, the expiratory intercostals, and the triangularis sterni, have a motor innervation that depends on segments situated in the thoracic or lum bar cord.I-3 Consequently, subjects with tetraplegia caused by transection of the cervical cord have severely compromised expiratory muscle function. Cough in these subjects is markedly impaired, and the clearance of bronchial secretions is defective. As a result, retention of secretions leading to atelectasis and bronchopul monary infections is a frequent occurrence. Expiratory muscle function, in fact, has traditionally been perceived as being totally lost in tetraplegia. Hence, cough would be a passive process resulting only from the elastic recoil properties of the respiratory system.4 Expiratory muscle function is totally lost indeed in subjects with transection of the upper cervical cord (CI-C4). However, a number of spirographic studies have established that most C5-C8 tetraplegic subjects generate a small expiratory reserve volume.5-1O Thus, although all the well-known muscles of expiration are paralysed, most tetraplegic subjects are still able to activate some muscles that can overcome the elastic resist ance to deflation of the chest wall and cause emptying of the lungs. If these muscles could be identified, one could train them for .strength and perhaps· improve the effectiveness of cough so as to reduce the number of bronchopulmonary infections. -

Svaly Hrudníku , Přehled Zádových Svalů
Muscles of thorax and abdomen. Muscle groups of the back. Vessels and nerves of the abdominal wall • Ivo Klepáček muscles inguina mm. epaxiales hypaxiales autochtonní heterochtonní musclesautochtonic heterochtonic Ep inguina Hyp Thoracic muscles • M.pectoralis major n.pectoralismuscles lat.+med. • M.pectoralis minor n.pectoralis med. • M.subclavius n.subclavius • M.serratus anterior n.thoracicus longus • Mm.intercostalesinguina externi, interni, intimi nn.intercostales I-XI • M.transversus thoracis nn.intercostales • Musculus diaphragma n.phrenicus M.pectoralis major Clavicular part Innervation: Sternal part pectoral nerves C5-Th1 Abdominalmuscles part adduction, rotation, accessory inspiratoryinguina muscle Muscles of the anterior axillar fold Thoracohumeral system M.pectoralis musclesminor inguina What compress subclavian or axillary artery A) Costa cervicalis B) mm. scaleni (m.scalenus minimus) C) Tumor inside spatium costoclaviculare D) Insertion of the m.pectoralis minor Intercostal muscles external intercostal m. M. intercostalis externus membrana intercostalis externa fascia thoracicamuscles superficialis (externa) internal intercostal m. M. intercostalis internus membrana intercostalis interna innermost intercostal m. M. intercostalis intimus fascia intercostalisinguina interna (endothoracica) Transverse thoracic m. M. transversus thoracis fascia intercostalis interna (endothoracica) Internal intercostal muscles + transversus ventral thoracic wall (dorsal view) muscles inguina m. transversus thoracis Internal and external -

Spontaneous Intercostal Bulge
Self-assessment corner 179 Postgrad Med J: first published as 10.1136/pgmj.74.869.179 on 1 March 1998. Downloaded from Spontaneous intercostal bulge Rodney J Folz, Claude A Piantadosi, Emily K Folz A 48-year-old man had been diagnosed with severe chronic obstructive pulmonary disease and referred to our pulmonary clinic. His medical history revealed a 60-80 pack year smoking history and long-standing heavy alcohol consumption. He worked as an upholsterer, with exposure to glues and paints for 20 years. Tuberculin skin testing was negative with a positive Candida con- trol. Sputum Gram-stain revealed many polymorphonuclear leukocytes and was AFB-stain posi- tive. Culture growth indicated mycobacterium other than tuberculosis identified as Runyoun's Group III. Drug sensitivity testing demonstrated 0% resistance to the combination of isoniazid, rifampin, and ethionamide. His clinical condition gradually deteriorated despite antimicrobial therapy. Pulmonary function testing in 1994 demonstrated severe obstructive lung disease with a DLCO of 29% predicted (8.5 ml/min/mmHg). One month later, he developed acute bronchitis and during a severe coughing spell he noticed the acute onset of bulging of his left anterior chest wall. The bulge rapidly expanded during manoeuvres which increased intrathoracic pressure and rapidly reduced with cessation of these manoeuvres. In addition to the expansion and contraction of this chest wall bulge, physical examination was remarkable for sonorous breath sounds and the sound of 'gurgling' fluid within the bulge. A chest X-ray is shown in figure 1. A chest computed tomogra- phy (CT) scan (figure 2) was obtained in the semi-prone position during a Valsalva manoeuvre. -

Acute Effects of the 6-Minute Pegboard and Ring Test in COPD
Acute Effects of the 6-Minute Pegboard and Ring Test in COPD Ivanize Mariana Masselli dos Reis, Renata Pedrolongo Basso-Vanelli, Thomas Beltrame, Maria Cecília Moraes Frade, Raphael Martins de Abreu, Marina Machado Cid, Aparecida Maria Catai, Ana Beatriz Oliveira, and Mauricio Jamami BACKGROUND: There are few reports in the literature supporting the understanding of the physiological mechanisms of intolerance in patients with COPD to perform unsupported upper limb activities. The aims of this study were to quantify the electrical activity and oxygenation of inspira- tory and upper limb muscles, and to investigate whether electromyographic manifestations of muscle fatigue are related to upper limb function as assessed by the 6-min pegboard and ring test (6PBRT) in subjects with COPD and in healthy subjects. METHODS: Thirty subjects with COPD ؎ ؎ (FEV1 42.1 16.4% predicted; 68.0 7.6 y old) comprised the COPD group, and 34 healthy subjects (66.8 ؎ 8.0 y old) comprised the control group. Both groups were assessed for body composition with dual-energy radiograph absorptiometry and spirometry. The 6PBRT was per- formed with simultaneous assessment of electromyography, near-infrared spectroscopy, and gas analyses (expiratory minute volume). RESULTS: Differences were observed between groups for performance (number of rings) in the 6PBRT, with the COPD group achieving lower values than the control group (P < .001). The ventilatory demand (expiratory minute volume/maximum vol- untary ventilation) and root mean square amplitude of the sternocleidomastoid muscle were higher in the COPD group than in the control group (P < .04). Lower values for oxyhemoglobin and total hemoglobin were found in intercostal muscles of the COPD group compared to the control group. -
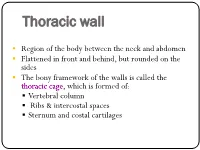
Thoracic Wall
Thoracic wall . Region of the body between the neck and abdomen . Flattened in front and behind, but rounded on the sides . The bony framework of the walls is called the thoracic cage, which is formed of: . Vertebral column . Ribs & intercostal spaces . Sternum and costal cartilages Superiorly: It communicates1st rib with the neck through an opening bounded:1 . Posteriorly by 1st thoracic vertebra . Laterally by medial border of the 1st ribs and their costal cartilages . Anteriorly by superior border of manubrium sterni Suprapleural This opening is occupied: membrane . In the midline, by the structures that pass between the neck and the thorax . On either sides, it is closed by a dense suprapleural membrane Suprapleural Membrane . Tent shaped dense fascial sheet that covers the apex of each lung. An extension of the endothoracic fascia . Extends approximately an inch superior to the superior thoracic aperture . It is attached: • The thoracic cage: . Protects the lungs, heart and large vessels . Provides attachment to the muscles of thorax, upper limb, abdomen & back • The cavity of thorax is divided into: • A median partition, the mediastinum • Laterally placed pleurae & lungs Cutaneous Nerves Anterior wall: . Above the level of sternal angle: Supraclavicular nerves . Below the level of sternal angle: Segmental innervation by anterior and lateral cutaneous branches of the intercostal nerves Posterior wall: . Segmental innervation by posterior rami of the thoracic spinal nerves nerves The Intercostal Space Intercostal Space It is the space between two ribs Since there are 12 ribs on each side, there are 11 intercostal spaces. Each space contains: . Intercostal muscles . Intercostal neurovascular bundle . Lymphatics Intercostal muscles • External Intercostal • Internal Intercostal • Innermost Intercostal Supplied by corresponding intercostal nerves Action: • Tend to pull the ribs nearer to each other .