04 34-35 Corticospinal Corticobulbar Pathways.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cranial Nerves
Cranial Nerves (1,5,7,8,9,10,11 and 12) Slides not included 9th and 10th Cranial 11th and 12th Cranial 8th Cranial Nerve 5th and 7th Cranial 1st Cranial Nerve Nerves Nerves Nerves (3,7,11,12,13,21,23,24) - (10,16) (12,23) Slides included: (14 to 17) *Slides that are not included mostly are slides of summaries or pictures. Nouf Alabdulkarim. Med 435 Olfactory Nerve [The 1st Cranial Nerve] Special Sensory Olfactory pathway 1st order neuron Receptors Axons of 1st order Neurons Olfactory receptors are specialized, ciliated nerve cells The axons of these bipolar cells 12 -20 fibers form the that lie in the olfactory epithelium. true olfactory nerve fibers. Which passes through the cribriform plate of ethmoid → They join the olfactory bulb Preliminary processing of olfactory information It is within the olfactory bulb, which contains interneurones and large Mitral cells; axons from the latter leave the bulb to form the olfactory tract. nd 2 order neuron • It is formed by the Mitral cells of olfactory bulb. • The axons of these cells form the olfactory tract. • Each tract divides into 2 roots at the anterior perforated substance: Lateral root Medial root Carries olfactory fibers to end in cortex of the Uncus & • crosses midline through anterior commissure adjacent part of Hippocampal gyrus (center of smell). and joins the uncrossed lateral root of opposite side. • It connects olfactory centers of 2 cerebral hemispheres. • So each olfactory center receives smell sensation from both halves of nasal cavity. NB. Olfactory pathway is the only sensory pathway which reaches the cerebral cortex without passing through the Thalamus . -

NS201C Anatomy 1: Sensory and Motor Systems
NS201C Anatomy 1: Sensory and Motor Systems 25th January 2017 Peter Ohara Department of Anatomy [email protected] The Subdivisions and Components of the Central Nervous System Axes and Anatomical Planes of Sections of the Human and Rat Brain Development of the neural tube 1 Dorsal and ventral cell groups Dermatomes and myotomes Neural crest derivatives: 1 Neural crest derivatives: 2 Development of the neural tube 2 Timing of development of the neural tube and its derivatives Timing of development of the neural tube and its derivatives Gestational Crown-rump Structure(s) age (Weeks) length (mm) 3 3 cerebral vesicles 4 4 Optic cup, otic placode (future internal ear) 5 6 cerebral vesicles, cranial nerve nuclei 6 12 Cranial and cervical flexures, rhombic lips (future cerebellum) 7 17 Thalamus, hypothalamus, internal capsule, basal ganglia Hippocampus, fornix, olfactory bulb, longitudinal fissure that 8 30 separates the hemispheres 10 53 First callosal fibers cross the midline, early cerebellum 12 80 Major expansion of the cerebral cortex 16 134 Olfactory connections established 20 185 Gyral and sulcul patterns of the cerebral cortex established Clinical case A 68 year old woman with hypertension and diabetes develops abrupt onset numbness and tingling on the right half of the face and head and the entire right hemitrunk, right arm and right leg. She does not experience any weakness or incoordination. Physical Examination: Vitals: T 37.0° C; BP 168/87; P 86; RR 16 Cardiovascular, pulmonary, and abdominal exam are within normal limits. Neurological Examination: Mental Status: Alert and oriented x 3, 3/3 recall in 3 minutes, language fluent. -

A Network of Genetic Repression and Derepression Specifies Projection
A network of genetic repression and derepression INAUGURAL ARTICLE specifies projection fates in the developing neocortex Karpagam Srinivasana,1, Dino P. Leonea, Rosalie K. Batesona, Gergana Dobrevab, Yoshinori Kohwic, Terumi Kohwi-Shigematsuc, Rudolf Grosschedlb, and Susan K. McConnella,2 aDepartment of Biology, Stanford University, Stanford, CA 94305; bMax-Planck Institute of Immunobiology and Epigenetics, 79108 Freiburg, Germany; and cLawrence Berkeley National Laboratory, Berkeley, CA 94720 This contribution is part of the special series of Inaugural Articles by members of the National Academy of Sciences elected in 2011. Contributed by Susan K. McConnell, September 28, 2012 (sent for review August 24, 2012) Neurons within each layer in the mammalian cortex have stereotypic which results in a suppression of corticothalamic and callosal projections. Four genes—Fezf2, Ctip2, Tbr1, and Satb2—regulate fates, respectively, and axon extension along the corticospinal these projection identities. These genes also interact with each tract (CST). Fezf2 mutant neurons fail to repress Satb2 and Tbr1, other, and it is unclear how these interactions shape the final pro- thus their axons cross the CC and/or innervate the thalamus jection identity. Here we show, by generating double mutants of inappropriately. Fezf2, Ctip2, and Satb2, that cortical neurons deploy a complex In callosal projection neurons, Satb2 represses the expression Ctip2 Bhlhb5 genetic switch that uses mutual repression to produce subcortical of and , leading to a repression of subcerebral fates. Satb2 Tbr1 or callosal projections. We discovered that Tbr1, EphA4, and Unc5H3 Interestingly, we found that promotes expression in fi upper layer callosal neurons, and Tbr1 expression in these neu- are critical downstream targets of Satb2 in callosal fate speci ca- fi tion. -
Corticospinal Fibers
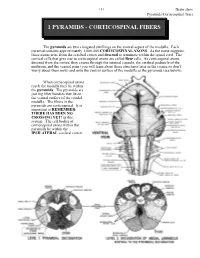
151 Brain stem Pyramids/Corticospinal Tract 1 PYRAMIDS - CORTICOSPINAL FIBERS The pyramids are two elongated swellings on the ventral aspect of the medulla. Each pyramid contains approximately 1,000,000 CORTICOSPINAL AXONS. As the name suggests, these axons arise from the cerebral cortex and descend to terminate within the spinal cord. The cortical cells that give rise to corticospinal axons are called Betz cells. As corticospinal axons descend from the cortex, they course through the internal capsule, the cerebral peduncle of the midbrain, and the ventral pons (you will learn about these structures later in the course so don’t worry about them now) and onto the ventral surface of the medulla as the pyramids (see below). When corticospinal axons reach the medulla they lie within the pyramids. The pyramids are just big fiber bundles that lie on the ventral surface of the caudal medulla. The fibers in the pyramids are corticospinal. It is important to REMEMBER: THERE HAS BEEN NO CROSSING YET! in this system. The cell bodies of corticospinal axons within the pyramids lie within the IPSILATERAL cerebral cortex. Brain stem 152 Pyramids/Corticospinal Tract At the most caudal pole of the pyramids the corticospinal axons cross over the midline and now continue their descent on the contralateral (to the cell of origin) side. This crossover point is called the PYRAMIDAL DECUSSATION. The crossing fibers enter the lateral funiculus of the spinal cord where they are called the LATERAL CORTICOSPINAL TRACT (“corticospinal” is not good enough, you have to call them lateral corticospinal; LCST - remember this one??). LCST axons exit the tract to terminate upon neurons in the spinal cord gray matter along its entire length. -

Projections from the Trigeminal Nuclear Complex to the Cochlear Nuclei: a Retrograde and Anterograde Tracing Study in the Guinea Pig
Journal of Neuroscience Research 78:901–907 (2004) Projections From the Trigeminal Nuclear Complex to the Cochlear Nuclei: a Retrograde and Anterograde Tracing Study in the Guinea Pig Jianxun Zhou and Susan Shore* Department of Otolaryngology and Kresge Hearing Research Institute, University of Michigan, Ann Arbor, Michigan In addition to input from auditory centers, the cochlear cuneate nucleus innervation of cochlear nucleus has been nucleus (CN) receives inputs from nonauditory centers, hypothesized to convey information about head and pinna including the trigeminal sensory complex. The detailed position for the purpose of localizing a sound source in anatomy, however, and the functional implications of the space (Young et al., 1995). In addition, interactions be- nonauditory innervation of the auditory system are not tween somatosensory and auditory systems have been fully understood. We demonstrated previously that the linked increasingly to phantom sound perception, also trigeminal ganglion projects to CN, with terminal labeling known as tinnitus. This is demonstrated in the observa- most dense in the marginal cell area and secondarily in tions that injuries of the head and neck region can lead to the magnocellular area of the ventral cochlear nucleus the onset of tinnitus in patients with no hearing loss (VCN). We continue this line of study by investigating the (Lockwood et al., 1998). projection from the spinal trigeminal nucleus to CN in We demonstrated previously projections from the guinea pig. After injections of the retrograde tracers Flu- trigeminal ganglion to CN in guinea pigs (Shore et al., oroGold or biotinylated dextran amine (BDA) in VCN, 2000). Terminal labeling of trigeminal ganglion projec- labeled cells were found in the spinal trigeminal nuclei, tions to the CN was found to be most dense in the most densely in the pars interpolaris and pars caudalis marginal cell area and secondarily in the magnocellular with ipsilateral dominance. -

The Superior and Inferior Colliculi of the Mole (Scalopus Aquaticus Machxinus)
THE SUPERIOR AND INFERIOR COLLICULI OF THE MOLE (SCALOPUS AQUATICUS MACHXINUS) THOMAS N. JOHNSON' Laboratory of Comparative Neurology, Departmmt of Amtomy, Un&versity of hfiehigan, Ann Arbor INTRODUCTION This investigation is a study of the afferent and efferent connections of the tectum of the midbrain in the mole (Scalo- pus aquaticus machrinus). An attempt is made to correlate these findings with the known habits of the animal. A subterranean animal of the middle western portion of the United States, Scalopus aquaticus machrinus is the largest of the genus Scalopus and its habits have been more thor- oughly studied than those of others of this genus according to Jackson ('15) and Hamilton ('43). This animal prefers a well-drained, loose soil. It usually frequents open fields and pastures but also is found in thin woods and meadows. Following a rain, new superficial burrows just below the surface of the ground are pushed in all directions to facili- tate the capture of worms and other soil life. Ten inches or more below the surface the regular permanent highway is constructed; the mole retreats here during long periods of dry weather or when frost is in the ground. The principal food is earthworms although, under some circumstances, larvae and adult insects are the more usual fare. It has been demonstrated conclusively that, under normal conditions, moles will eat vegetable matter. It seems not improbable that they may take considerable quantities of it at times. A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the University of Michigan. -

The Use of Cholinesterase Techniques to Study Topographical Localization in the Hypoglossal Nucleus of the Rat
J. Anat. (1971), 110, 2, pp. 203-213 203 With 1O figures Printed in Great Britain The use of cholinesterase techniques to study topographical localization in the hypoglossal nucleus of the rat P. R. LEWIS*, B. A. FLUMERFELTt AND C. C. D. SHUTE* Department ofAnatomy, University of Cambridge (Accepted 4 August 1971) INTRODUCTION The hypoglossal nucleus is perhaps the most suitable motor nucleus for the experi- mental study of the cytological changes occurring in cholinergic neurons following axotomy. The cells are large and the nucleus is easy to find even in a fresh unfixed brain; furthermore, the nucleus is so close to the midline that it is possible to use one side as a control for the other with complete confidence and to view equivalent control and experimental neurons simultaneously at quite high magnifications. An added advantage is that the hypoglossal nerve trunk in the neck region is almost purely motor; the central effects of axotomy are therefore not complicated by any significant loss of sensory fibres. Our interest in the nucleus was heightened by the discovery that in the rat a group of neurons at the caudal end contained a high concentration of an enzyme resembling in its histochemical reactions pseudocholin- esterase (Shute & Lewis, 1963). The enzyme will hydrolyse acetylthiocholine and is inhibited by ethopropazine, but its most characteristic property is a rapid hydrolysis of butyrylthiocholine; BuChE would thus seem an appropriate abbreviation to distinguish it from true cholinesterase (AChE), the enzyme typically present in motor neurons. It was shown originally by Schwarzacher (1958) that there is a marked decrease in AChE activity in hypoglossal neurons during the second and third weeks following axotomy (although he also looked at the response of pseudocholinesterase he did not comment on the specifically staining group of cells). -

Circuits in the Rodent Brainstem That Control Whisking in Concert with Other Orofacial Motor Actions
Neuroscience 368 (2018) 152–170 CIRCUITS IN THE RODENT BRAINSTEM THAT CONTROL WHISKING IN CONCERT WITH OTHER OROFACIAL MOTOR ACTIONS y y LAUREN E. MCELVAIN, a BETH FRIEDMAN, a provides the reset to the relevant premotor oscillators. HARVEY J. KARTEN, b KAREL SVOBODA, c FAN WANG, d Third, direct feedback from somatosensory trigeminal e a,f,g MARTIN DESCHEˆ NES AND DAVID KLEINFELD * nuclei can rapidly alter motion of the sensors. This feed- a Department of Physics, University of California at San Diego, back is disynaptic and can be tuned by high-level inputs. La Jolla, CA 92093, USA A holistic model for the coordination of orofacial motor actions into behaviors will encompass feedback pathways b Department of Neurosciences, University of California at San Diego School of Medicine, La Jolla, CA 92093, USA through the midbrain and forebrain, as well as hindbrain c areas. Howard Hughes Medical Institute, Janelia Research This article is part of a Special Issue entitled: Barrel Campus, Ashburn, VA 20147, USA Cortex. Ó 2017 IBRO. Published by Elsevier Ltd. All rights d Department of Neurobiology, Duke University Medical reserved. Center, Durham, NC 27710, USA e Department of Psychiatry and Neuroscience, Laval University, Que´bec City, G1J 2G3, Canada f Key words: coupled oscillators, facial nucleus, hypoglossal Section of Neurobiology, University of California at San Diego, La Jolla, CA 92093, USA nucleus, licking, orienting, tongue, vibrissa. g Department of Electrical and Computer Engineering, University of California at San Diego, La Jolla, CA 92093, USA Contents Introduction 153 Abstract—The world view of rodents is largely determined Coordination of multiple orofacial motor actions 153 by sensation on two length scales. -

The Brain Stem Medulla Oblongata
Chapter 14 The Brain Stem Medulla Oblongata Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Central sulcus Parietal lobe • embryonic myelencephalon becomes Cingulate gyrus leaves medulla oblongata Corpus callosum Parieto–occipital sulcus Frontal lobe Occipital lobe • begins at foramen magnum of the skull Thalamus Habenula Anterior Epithalamus commissure Pineal gland • extends for about 3 cm rostrally and ends Hypothalamus Posterior commissure at a groove between the medulla and Optic chiasm Mammillary body pons Cerebral aqueduct Pituitary gland Fourth ventricle Temporal lobe • slightly wider than spinal cord Cerebellum Midbrain • pyramids – pair of external ridges on Pons Medulla anterior surface oblongata – resembles side-by-side baseball bats (a) • olive – a prominent bulge lateral to each pyramid • posteriorly, gracile and cuneate fasciculi of the spinal cord continue as two pair of ridges on the medulla • all nerve fibers connecting the brain to the spinal cord pass through the medulla • four pairs of cranial nerves begin or end in medulla - IX, X, XI, XII Medulla Oblongata Associated Functions • cardiac center – adjusts rate and force of heart • vasomotor center – adjusts blood vessel diameter • respiratory centers – control rate and depth of breathing • reflex centers – for coughing, sneezing, gagging, swallowing, vomiting, salivation, sweating, movements of tongue and head Medulla Oblongata Nucleus of hypoglossal nerve Fourth ventricle Gracile nucleus Nucleus of Cuneate nucleus vagus -

Imaging of the Confused Patient: Toxic Metabolic Disorders Dara G
Imaging of the Confused Patient: Toxic Metabolic Disorders Dara G. Jamieson, M.D. Weill Cornell Medicine, New York, NY The patient who presents with either acute or subacute confusion, in the absence of a clearly defined speech disorder and focality on neurological examination that would indicate an underlying mass lesion, needs to be evaluated for a multitude of neurological conditions. Many of the conditions that produce the recent onset of alteration in mental status, that ranges from mild confusion to florid delirium, may be due to infectious or inflammatory conditions that warrant acute intervention such as antimicrobial drugs, steroids or plasma exchange. However, some patients with recent onset of confusion have an underlying toxic-metabolic disorders indicating a specific diagnosis with need for appropriate treatment. The clinical presentations of some patients may indicate the diagnosis (e.g. hypoglycemia, chronic alcoholism) while the imaging patterns must be recognized to make the diagnosis in other patients. Toxic-metabolic disorders constitute a group of diseases and syndromes with diverse causes and clinical presentations. Many toxic-metabolic disorders have no specific neuroimaging correlates, either at early clinical stages or when florid symptoms develop. However, some toxic-metabolic disorders have characteristic abnormalities on neuroimaging, as certain areas of the central nervous system appear particularly vulnerable to specific toxins and metabolic perturbations. Areas of particular vulnerability in the brain include: 1) areas of high-oxygen demand (e.g. basal ganglia, cerebellum, hippocampus), 2) the cerebral white matter and 3) the mid-brain. Brain areas of high-oxygen demand are particularly vulnerable to toxins that interfere with cellular respiratory metabolism. -

Efficacy And/Or Effectiveness of Portable Neuromodulation Stimulator (Pons®) As Treatment for Traumatic Brain Injury (TBI)”
Evidence-Based Practice Group Answers to Clinical Questions “Efficacy and/or Effectiveness of Portable Neuromodulation Stimulator (PoNS®) as Treatment for Traumatic Brain Injury (TBI)” A Rapid Systematic Review By WorkSafeBC Evidence-Based Practice Group Dr. Craig Martin Manager, Clinical Services Chair, Evidence-Based Practice Group October 2019 Clinical Services – Worker and Employer Services Efficacy and/or Effectiveness of Portable Neuromodulation Stimulator (PoNS®) as Treatment for Traumatic Brain Injury (TBI) i About this report Efficacy and/or Effectiveness of Portable Neuromodulation Stimulator (PoNS®) as Treatment for Traumatic Brain Injury (TBI) Published: October 2019 About the Evidence-Based Practice Group The Evidence-Based Practice Group was established to address the many medical and policy issues that WorkSafeBC officers deal with on a regular basis. Members apply established techniques of critical appraisal and evidence-based review of topics solicited from both WorkSafeBC staff and other interested parties such as surgeons, medical specialists, and rehabilitation providers. Suggested Citation WorkSafeBC Evidence-Based Practice Group, Martin CW. Efficacy and/or Effectiveness of Portable Neuromodulation Stimulator (PoNS®) as Treatment for Traumatic Brain Injury (TBI). Richmond, BC: WorksafeBC Evidence- Based Practice Group; October 2019. Contact Information Evidence-Based Practice Group WorkSafeBC PO Box 5350 Stn Terminal Vancouver BC V6B 5L5 Email [email protected] Phone 604 279-7417 Toll-free 1 888 967-5377 -

DR. Sanaa Alshaarawy
By DR. Sanaa Alshaarawy 1 By the end of the lecture, students will be able to : Distinguish the internal structure of the components of the brain stem in different levels and the specific criteria of each level. 1. Medulla oblongata (closed, mid and open medulla) 2. Pons (caudal, mid “Trigeminal level” and rostral). 3. Mid brain ( superior and inferior colliculi). Describe the Reticular formation (structure, function and pathway) being an important content of the brain stem. 2 1. Traversed by the Central Canal. Motor Decussation*. Spinal Nucleus of Trigeminal (Trigeminal sensory nucleus)* : ➢ It is a larger sensory T.S of Caudal part of M.O. nucleus. ➢ It is the brain stem continuation of the Substantia Gelatinosa of spinal cord 3 The Nucleus Extends : Through the whole length of the brain stem and upper segments of spinal cord. It lies in all levels of M.O, medial to the spinal tract of the trigeminal. It receives pain and temperature from face, forehead. Its tract present in all levels of M.O. is formed of descending fibers that terminate in the trigeminal nucleus. 4 It is Motor Decussation. Formed by pyramidal fibers, (75-90%) cross to the opposite side They descend in the Decuss- = crossing lateral white column of the spinal cord as the lateral corticospinal tract. The uncrossed fibers form the ventral corticospinal tract. 5 Traversed by Central Canal. Larger size Gracile & Cuneate nuclei, concerned with proprioceptive deep sensations of the body. Axons of Gracile & Cuneate nuclei form the internal arcuate fibers; decussating forming Sensory Decussation. Pyramids are prominent ventrally. 6 Formed by the crossed internal arcuate fibers Medial Leminiscus: Composed of the ascending internal arcuate fibers after their crossing.