Research Commons at The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hamilton Gardens Waikato Museum
‘Shovel ready’ Infrastructure Projects: Project Information Form About this Project Information Form The Government is seeking to identify ‘shovel ready’ infrastructure projects from the Public and certain Private Infrastructure sector participants that have been impacted by COVID 19. Ministers have advised that they wish to understand the availability, benefits, geographical spread and scale of ‘shovel ready’ projects in New Zealand. These projects will be considered in the context of any potential Government response to support the construction industry, and to provide certainty on a pipeline of projects to be commenced or re- commenced, once the COVID 19 Response Level is suitable for construction to proceed. The Infrastructure Industry Reference Group, chaired by Mark Binns, is leading this work at the request of Ministers, and is supported by Crown Infrastructure Partners Limited (CIP). CIP is now seeking information using this Project Information Form from relevant industry participants for 1 projects/programmes that may be suitable for potential Government support. The types of projects we have been asked to consider is outlined in Mark Binns’ letter dated 25 March 2020. CIP has prepared Project Information Guidelines which outline the approach CIP will take in reviewing and categorising the project information it receives (Guidelines). Please submit one form for each project that you consider meets the criteria set out in the Guidelines. If you have previously provided this information in another format and/or as part of a previous process feel free to submit it in that format and provide cross-references in this form. Please provide this information by 5 pm on Tuesday 14 April 2020. -
Attachment 4
PARK ROAD SULLIVAN ROAD DIVERS ROAD SH 1 LAW CRESCENT BIRDWOOD ROAD WASHER ROAD HOROTIU BRIDGE ROAD CLOVERFIELD LANE HOROTIU ROAD KERNOTT ROAD PATERSON ROAD GATEWAY DRIVE EVOLUTION DRIVE HE REFORD D RIVE IN NOVA TIO N W A Y MARTIN LANE BOYD ROAD HENDERSON ROAD HURRELL ROAD HUTCHINSON ROAD BERN ROAD BALLARD ROAD T NNA COU R VA SA OSBORNE ROAD RK D RIVER DOWNS PA RIVE E WILLIAMSON ROAD G D RI ONION ROAD C OU N T REYNOLDS ROAD RY L ANE T E LANDON LANE R A P A A ROAD CESS MEADOW VIEW LANE Attachment 4 C SHERWOOD DOWNS DRIVE HANCOCK ROAD KAY ROAD REDOAKS CLOSE REID ROAD RE E C SC E V E EN RI L T IV D A R GRANTHAM LANE KE D D RIVERLINKS LANE RIVERLINKS S K U I P C A E L O M C R G N E A A A H H D O LOFTUS PLACE W NORTH CITY ROAD F I ONION ROAD E LD DROWERGLEN S Pukete Farm Park T R R O SE EE T BE MCKEE STREET R RY KUPE PLACE C LIMBER HILL HIGHVIEW COURT RESCENT KESTON CRESCENT VIKING LANE CLEWER LANE NICKS WAY CUM BE TRAUZER PLACE GRAHAM ROAD OLD RUFFELL ROAD RLA KOURA DRIVE ND DELIA COURT DR IV ET ARIE LANE ARIE E E SYLVESTER ROAD TR BREE PLACE E S V TRENT LANE M RI H A D ES G H UR C B WESSEX PLACE WALTHAM PLACE S GO R IA P Pukete Farm Park T L T W E M A U N CE T I HECTOR DRIVE I A R H E R S P S M A AMARIL LANE IT IR A N M A N I N G E F U A E G ED BELLONA PLACE Y AR D M E P R S T L I O L IN Moonlight Reserve MAUI STREET A I A L G C A P S D C G A D R T E RUFFELL ROAD R L L A O E O N R A E AVALON DRIVEAVALON IS C N S R A E R E D C Y E E IS E E L C N ANN MICHELE STREET R E A NT ESCE NT T WAKEFIELD PLACE E T ET B T V TE KOWHAI ROAD KAPUNI STRE -

Plant Charts for Native to the West Booklet
26 Pohutukawa • Oi exposed coastal ecosystem KEY ♥ Nurse plant ■ Main component ✤ rare ✖ toxic to toddlers coastal sites For restoration, in this habitat: ••• plant liberally •• plant generally • plant sparingly Recommended planting sites Back Boggy Escarp- Sharp Steep Valley Broad Gentle Alluvial Dunes Area ment Ridge Slope Bottom Ridge Slope Flat/Tce Medium trees Beilschmiedia tarairi taraire ✤ ■ •• Corynocarpus laevigatus karaka ✖■ •••• Kunzea ericoides kanuka ♥■ •• ••• ••• ••• ••• ••• ••• Metrosideros excelsa pohutukawa ♥■ ••••• • •• •• Small trees, large shrubs Coprosma lucida shining karamu ♥ ■ •• ••• ••• •• •• Coprosma macrocarpa coastal karamu ♥ ■ •• •• •• •••• Coprosma robusta karamu ♥ ■ •••••• Cordyline australis ti kouka, cabbage tree ♥ ■ • •• •• • •• •••• Dodonaea viscosa akeake ■ •••• Entelea arborescens whau ♥ ■ ••••• Geniostoma rupestre hangehange ♥■ •• • •• •• •• •• •• Leptospermum scoparium manuka ♥■ •• •• • ••• ••• ••• ••• ••• ••• Leucopogon fasciculatus mingimingi • •• ••• ••• • •• •• • Macropiper excelsum kawakawa ♥■ •••• •••• ••• Melicope ternata wharangi ■ •••••• Melicytus ramiflorus mahoe • ••• •• • •• ••• Myoporum laetum ngaio ✖ ■ •••••• Olearia furfuracea akepiro • ••• ••• •• •• Pittosporum crassifolium karo ■ •• •••• ••• Pittosporum ellipticum •• •• Pseudopanax lessonii houpara ■ ecosystem one •••••• Rhopalostylis sapida nikau ■ • •• • •• Sophora fulvida west coast kowhai ✖■ •• •• Shrubs and flax-like plants Coprosma crassifolia stiff-stemmed coprosma ♥■ •• ••••• Coprosma repens taupata ♥ ■ •• •••• •• -

Brymer Crawshaw Nawton Pukete Flagstaff Sylvester Maeroa Bryant
M R T H A I U V E I E T L S E IN O T C R N K R A R E N E L C ROV E P H G E N O S Y I T A A E E O AN N E E R H D S R S L E T C L V I GAL N E C I E Y E R R V S R I R E Guide to using this map: D R L O A L D Y I A E P E H N E R A E C D A L C E G C E A A A D D R I I V L E S P This is a map of the area containing your property. R E O E C O L R D I C S I C E A A M O N P I V T R L R I H O S L E D E O I T R W R R C L D E H O D C K P M O O S V H O R E S R AVE P A G A VOU NUE E NT N A W C A The map shows notable local changes which are C R E S CE T Y DE R N B E EET R R I E ST VE proposed for the mapped area. E C E L T R IC H E E L A M R R Z N T O IM A N I S A I N U U D P T T P Sylvester H KA TA S H K E P E L S R I A B A S C E E See the map legend for an index of these local O N U P L L E A changes and check the map to see which ones V E C C D A LA R P E T P A E Flagstaff Y S E E U O D IM R U E affect your area of the city. -

8 February 2012 Time: 9.30 Am Meeting Room: Committee Room 1 Venue: Municipal Building, Garden Place, Hamilton
Notice of Meeting: I hereby give notice that an ordinary meeting of Operations & Activty Performance Committee will be held on: Date: Wednesday, 8 February 2012 Time: 9.30 am Meeting Room: Committee Room 1 Venue: Municipal Building, Garden Place, Hamilton Barry Harris Chief Executive Operations & Activity Performance Committee OPEN AGENDA Membership Chairperson Cr M Gallagher Deputy Chairperson Cr A O’Leary Members Her Worship the Mayor Ms J Hardaker Cr D Bell Cr P Bos Cr G Chesterman Cr M Forsyth Cr J Gower Cr R Hennebry Cr D Macpherson Cr P Mahood Cr M Westphal Cr E Wilson Quorum: A majority of members (including vacancies) Meeting Frequency: Monthly Fleur Yates Senior Committee Advisor 1 February 2012 [email protected] Telephone: 838 6771 www.hamilton.co.nz Operations & Activity Performance Committee Agenda 8 February 2012- OPEN Page 1 of 145 Role & Scope . The overall mandate of this committee is to request and receive information concerning Councils activities and develop consistent and pragmatic reasoning that will enable Council to be informed of future directions, options and choices. The committee has no decision making powers unless for minor matters that improve operational effectiveness, efficiency or economy. To monitor key activities and services (without operational interference in the services) in order to better inform elected members and the community about key Council activities and issues that arise in the operational arm of the Council. No more than 2 operational areas to report each month . Receive reports relating to organisational performance against KPI’s, delivery of strategic goals, and community outcomes and vision. -

Pteridologist 2007
PTERIDOLOGIST 2007 CONTENTS Volume 4 Part 6, 2007 EDITORIAL James Merryweather Instructions to authors NEWS & COMMENT Dr Trevor Walker Chris Page 166 A Chilli Fern? Graham Ackers 168 The Botanical Research Fund 168 Miscellany 169 IDENTIFICATION Male Ferns 2007 James Merryweather 172 TREE-FERN NEWSLETTER No. 13 Hyper-Enthusiastic Rooting of a Dicksonia Andrew Leonard 178 Most Northerly, Outdoor Tree Ferns Alastair C. Wardlaw 178 Dicksonia x lathamii A.R. Busby 179 Tree Ferns at Kells House Garden Martin Rickard 181 FOCUS ON FERNERIES Renovated Palace for Dicksoniaceae Alastair C. Wardlaw 184 The Oldest Fernery? Martin Rickard 185 Benmore Fernery James Merryweather 186 FEATURES Recording Ferns part 3 Chris Page 188 Fern Sticks Yvonne Golding 190 The Stansfield Memorial Medal A.R. Busby 191 Fern Collections in Manchester Museum Barbara Porter 193 What’s Dutch about Dutch Rush? Wim de Winter 195 The Fine Ferns of Flora Græca Graham Ackers 203 CONSERVATION A Case for Ex Situ Conservation? Alastair C. Wardlaw 197 IN THE GARDEN The ‘Acutilobum’ Saga Robert Sykes 199 BOOK REVIEWS Encyclopedia of Garden Ferns by Sue Olsen Graham Ackers 170 Fern Books Before 1900 by Hall & Rickard Clive Jermy 172 Britsh Ferns DVD by James Merryweather Graham Ackers 187 COVER PICTURE: The ancestor common to all British male ferns, the mountain male fern Dryopteris oreades, growing on a ledge high on the south wall of Bealach na Ba (the pass of the cattle) Unless stated otherwise, between Kishorn and Applecross in photographs were supplied the Scottish Highlands - page 172. by the authors of the articles PHOTO: JAMES MERRYWEATHER in which they appear. -

Peachgrove Maeroa Swarbrick Clarkin Hamilton East Chartwell
T P E D B A A U A A R L G O W T A N RFIELD C E A AI R R P R A B F E E R O S A A R C S T U R N D D E T T R I E O N S E E O R T T S A R T E Y ESCE T S R NT D C R C L M E S Guide to using this map: E A H E C AT V R R A C E F R Y I I V E S R A H R D E T I E N O E E A S IE R D E V U D N S OA E S A R A Y E I V K S P T D C U C C P L R O C A This is a map of the area containing your property. E A E H C E E T E R Y E E T M N S U D N R T T L E W P O O T E S R R E A A U O R T A L A E N D D A T E E C C A H W A N C L S V E AL T O R V E SH R H A A O N F I R R L R C D A E The map shows notable local changes which are Y S U B A M G B T R S O P E I O A R S E O R R S T R R U L D T Y I N T A L L O O R N A R A T M S O L O L Y L E M S E T C O V IT O N R A H proposed for the mapped area. -

Minutes of Ordinary Council Meeting
Council 14 MAY 2018 - OPEN Council 10 Year Plan Hearings OPEN MINUTES Minutes of meetings of the Council held in the Council Chamber, Municipal Building, Garden Place, Hamilton on Friday 11 May 2018 at 9.40am and Monday 14 May 2018 (which reconvened Tuesday - Thursday 15-17 May 2018). The reports for both these meetings were contained within the agenda of the Extraordinary Council meeting of 11 May 2018. PRESENT Chairperson Mayor A King Deputy Chairperson Deputy Mayor M Gallagher Members Cr M Bunting Cr J R Casson Cr S Henry Cr D Macpherson Cr G Mallett Cr A O’Leary Cr R Pascoe Cr P Southgate Cr G Taylor Cr L Tooman Cr R Hamilton In Attendance: Richard Briggs – Chief Executive Lance Vervoort – General Manager Community Sean Hickey – General Manager Strategy and Communication David Bryant – General Manager Corporate Chris Allen - General Manager Infrastructure Jen Baird - General Manager City Growth Blair Bowcott – Executive Director Special Projects Julie Clausen – Programme Manager Chelsey Stewart – Project Manager 10 Year Plan Nigel Ward - Acting Communications Team Leader Andy Mannering – Manager Social Development Andrew Parsons - City Development Manager Greg Carstens – Acting Unit Manager Economic Growth & Planning Nathan Dalgety – Team Leader Growth Funding & Analytics Stafford Hodgson – Senior Strategic Policy Analyst Muna Wharawhara – Amorangi Maaori Governance Staff: Lee-Ann Jordan - Governance Manager Becca Brooke – Governance Team Leader Amy Viggers, Claire Guthrie and Rebecca Watson – Committee Advisor Muna Wharawhara carried out a blessing and Rev Phil Wilson a reading to open the Council Meeting. COUNCIL 14 MAY 2018 -OPEN Page 1 of 29 Council 14 MAY 2018 - OPEN 1. -

List of Road Names in Hamilton
Michelle van Straalen From: official information Sent: Monday, 3 August 2020 16:30 To: Cc: official information Subject: LGOIMA 20177 - List of road and street names in Hamilton. Attachments: FW: LGOIMA 20177 - List of road and street names in Hamilton. ; LGOIMA - 20177 Street Names.xlsx Kia ora Further to your information request of 6 July 2020 in respect of a list of road and street names in Hamilton, I am now able to provide Hamilton City Council’s response. You requested: Does the Council have a complete list of road and street names? Our response: Please efind th information you requested attached. We trust this information is of assistance to you. Please do not hesitate to contact me if you have any further queries. Kind regards, Michelle van Straalen Official Information Advisor | Legal Services | Governance Unit DDI: 07 974 0589 | [email protected] Hamilton City Council | Private Bag 3010 | Hamilton 3240 | www.hamilton.govt.nz Like us on Facebook Follow us on Twitter This email and any attachments are strictly confidential and may contain privileged information. If you are not the intended recipient please delete the message and notify the sender. You should not read, copy, use, change, alter, disclose or deal in any manner whatsoever with this email or its attachments without written authorisation from the originating sender. Hamilton City Council does not accept any liability whatsoever in connection with this email and any attachments including in connection with computer viruses, data corruption, delay, interruption, unauthorised access or unauthorised amendment. Unless expressly stated to the contrary the content of this email, or any attachment, shall not be considered as creating any binding legal obligation upon Hamilton City Council. -

Pteridologist 2009
PTERIDOLOGIST 2009 Contents: Volume 5 Part 2, 2009 The First Pteridologist Alec Greening 66 Back from the dead in Corrie Fee Heather McHaffie 67 Fern fads, fashions and other factors Alec Greening 68 A Stumpery on Vashon Island near Seattle Pat Reihl 71 Strange Revisions to The Junior Oxford English Dictionary Alistair Urquhart 73 Mauchline Fern Ware Jennifer Ide 74 More Ferns In Unusual Places Bryan Smith 78 The Ptéridophytes of Réunion Edmond Grangaud 79 Croziers - a photographic study. Linda Greening 84 A fern by any other name John Edgington 85 Tree-Fern Newsletter No. 15 Edited by Alastair C. Wardlaw 88 Editorial: TFNL then and now Alastair C. Wardlaw 88 Courtyard Haven for Tree Ferns Alastair C. Wardlaw 88 Bulbils on Tree Ferns: II Martin Rickard 90 Gough-Island Tree Fern Comes to Scotland Jamie Taggart 92 Growing ferns in a challenging climate Tim Pyner 95 Maraudering caterpillars. Yvonne Golding 104 New fern introductions from Fibrex Nurseries Angela Tandy 105 Ferns which live with ants! Yvonne Golding 108 The British Fern Gazette 1909 – 2009 Martin Rickard 110 A Siberian Summer Chris Page 111 Monitoring photographs of Woodsia ilvenis Heather McHaffie 115 Notes on Altaian ferns Irina Gureyeva 116 Ferns from the Galapagos Islands. Graham Ackers 118 Did you know? (Extracts from the first Pteridologist) Jimmy Dyce 121 The First Russian Pteridological Conference Chris Page 122 Tectaria Mystery Solved Pat Acock 124 Chatsworth - a surprising fern link with the past Bruce Brown 125 Fern Postage Stamps from the Faroe Islands Graham Ackers 127 Carrying out trials in your garden Yvonne Golding 128 A national collection of Asplenium scolopendrium Tim Brock 130 Asplenium scolopendrium ‘Drummondiae’ Tim Brock 132 Fern Recording – A Personal Scottish Experience Frank McGavigan 133 Book Notes Martin Rickard 136 Gay Horsetails Wim de Winter 137 Ferning in snow Martin Rickard 139 Fern Enthusiasts do the strangest things. -

Te Awa Lakes: Housing Economics
5 April 2018 Attn: Paul Radich Development Planner, Te Awa Lakes Development Perry Group Via email: [email protected] CC: [email protected] Te Awa Lakes: Housing Economics Dear Paul, This letter aims to provide ongoing assessment in relation to demand for Qualifying Developments, and also comments on local demand for residential housing in the proposed Te Awa Lakes Special Housing Area. Updating RCG’s Housing Commentary We summarise the key points from RCG’s Assessment of Economic Effects report for Te Awa Lakes below. 1 We have updated these points with more recent data where available: • Net migration into New Zealand remains at near-record levels of around 70,000 a year. • Auckland is still not keeping pace with its own housing shortage, i.e. ‘supply’ of new homes is not matching ‘demand’ from population growth. • Auckland is likely to keep losing people to neighbouring regions such as the Waikato, especially when there is housing pressure (as is currently the case). • Auckland’s house price boom began in 2012 and spread to Hamilton by 2015. In both cities, and most other parts of New Zealand, prices have flattened out in 2017-18. • The average house value in Hamilton is now $548,000, compared with $363,000 four years ago. 2 • Building consents in Hamilton, Waikato and Waipa remain at near-record levels, but have plateaued. 1 http://www.hamilton.govt.nz/our-council/council- publications/districtplans/ODP/Documents/Te%20Awa%20Lakes%20Private%20Plan%20Chan ge/Appendix_6_Assessment_of_Economic_Effects.PDF 2 Data from https://www.qv.co.nz/property-trends/residential-house-values , for Feb 2018 compared with Feb 2014 RCG | CONSTRUCTIVE THINKING. -
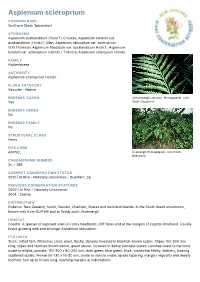
Asplenium Scleroprium
Asplenium scleroprium COMMON NAME Southern Shore Spleenwort SYNONYMS Asplenium aucklandicum (Hook.f.) Crookes; Asplenium lucidum var. aucklandicum (Hook.f.) Allan; Asplenium obtusatum var. scleroprium G.M.Thomson; Asplenium flaccidum var. aucklandicum Hook.f.; Asplenium lucidum var. scleroprium (Hombr.) T.Moore; Asplenium scleropium Hombr. FAMILY Aspleniaceae AUTHORITY Asplenium scleroprium Hombr. FLORA CATEGORY Vascular – Native ENDEMIC TAXON At Invercargill (January). Photographer: John Yes Smith-Dodsworth ENDEMIC GENUS No ENDEMIC FAMILY No STRUCTURAL CLASS Ferns NVS CODE ASPSCL Invercargill. Photographer: John Smith- Dodsworth CHROMOSOME NUMBER 2n = 288 CURRENT CONSERVATION STATUS 2012 | At Risk – Naturally Uncommon | Qualifiers: Sp PREVIOUS CONSERVATION STATUSES 2009 | At Risk – Naturally Uncommon 2004 | Sparse DISTRIBUTION Endemic. New Zealand, South, Stewart, Chatham, Snares and Auckland Islands. In the South Island uncommon, known only from Bluff Hill and at Sandy point, Invercargill. HABITAT Coastal. A species of exposed sites on rocky headlands, cliff faces and at the margins of coastal shrubland. Usually found growing with and amongst Asplenium obtusatum. FEATURES Stout, tufted fern. Rhizomes stout, erect, fleshy, densely invested in blackish-brown scales. Stipes 150-500 mm long, stipes and rachises brown below, green above. Covered in dense subulate scales. Laminae ovate to narrowly ovate or elliptic, pinnate, 150-500 x 80-200 mm, dark green, blue green, thick, somewhat fleshy, leathery, bearing scattered scales. Pinnae 50-130 x 10-20 mm, ovate to narrow ovate, apices tapering, margins regularly and deeply toothed. Sori up to 10 mm long, reaching margins at indentations SIMILAR TAXA Most likely to be confused with A. obtusatum with which it frequently grows and sometimes hybridises with.