Human Anatomy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Autologous Gluteal Lipograft
Aesth Plast Surg (2011) 35:216–224 DOI 10.1007/s00266-010-9590-y ORIGINAL ARTICLE Autologous Gluteal Lipograft Beatriz Nicareta • Luiz Haroldo Pereira • Aris Sterodimas • Yves Ge´rard Illouz Received: 14 January 2010 / Accepted: 15 July 2010 / Published online: 25 September 2010 Ó Springer Science+Business Media, LLC and International Society of Aesthetic Plastic Surgery 2010 Abstract In the past 25 years, several different tech- expressed the desire of further gluteal augmentation, 16 had niques of lipoinjection have been developed. The authors one more session of gluteal fat grafting. The remaining five performed a prospective study to evaluate the patient sat- patients did not have enough donor area and instead isfaction and the rate of complications after an autologous received gluteal silicone implants. At 12 months, 70% gluteal lipograft among 351 patients during January 2002 reported that their appearance after gluteal fat augmentation and January 2008. All the patients included in the study was ‘‘very good’’ to ‘‘excellent,’’ and 23% responded that requested gluteal augmentation and were candidates for their appearance was ‘‘good.’’ Only 7% of the patients the procedure. Overall satisfaction with body appearance thought their appearance was less than good. At 24 months, after gluteal fat augmentation was rated on a scale of 1 66% reported that their appearance after gluteal fat aug- (poor), 2 (fair), 3 (good), 4 (very good), and 5 (excellent). mentation was ‘‘very good’’ (36%) to ‘‘excellent’’ (30%), The evaluation was made at follow-up times of 12 and and 27% responded that their appearance was ‘‘good.’’ 24 months. The total amount of clean adipose tissue However, 7% of the patients continued to think that their transplanted to the buttocks varied from 100 to 900 ml. -

The Abdominal Wall the Digestive Tract the Pancreas the Biliary
The abstracts which follow have been classified for the convenience of the reader under the following headings: Experimental Studies; Animal Tumors The Abdominal Wall The Cancer Cell The Digestive Tract General Clinical and Laboratory Observa- The Pancreas tions The Biliary Tract Diagnosis and Treatment Peritoneal, Retroperitoneal. and Mesenteric The Skin Tumors The Eye The Spleen The Ear The Female Genital Tract The Breast The Genito-Urinary Tract The Oral Cavity and Upper Respiratory The Nervous System Tract The Bones and Joints The Salivary Glands The Leukemias, Hodgkin's Disease, Lympho The Thyroid Gland sarcoma Intrathoracic Tumors As with any such scheme of classification, overlapping has been unavoidable. Shall an article on II Cutaneous Melanoma, an Histological Study" be grouped with the articles on Histology or with the Skin Tumors? Shall Traumatic Cerebral Tumors go under Trauma or The Nervous System? The reader's choice is likely to depend upon his personal interests; an editor may be governed by no such considerations. The attempt has been made, there fore, to put such articles in the group where they would seem most likely to be sought by the greatest number. It is hoped that this aim has not been entirely missed. As abstractors are never perfect, and as the opinions expressed may on occasion seem to an author not to represent adequately his position, opportunity is offered any such to submit his own views for publication. The JOURNAL will not only welcome correspondence of this nature but hopes in the future to have a large number of author abstracts, so that the writer of a paper may present his subject in his own way. -
Pubid-1446077038.Pdf
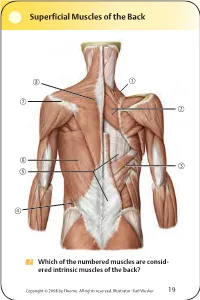
Superficial Muscles of the Back Which of the numbered muscles are consid- ered intrinsic muscles of the back? Copyright © 2008 by Thieme. All rights reserved. Illustrator: Karl Wesker 19 Superficial Muscles of the Back Posterior view. A Levator scapulae S Rhomboideus major D Serratus posterior inferior F Lumbar triangle, internal oblique G Thoracolumbar fascia, superficial layer H Latissimus dorsi J Scapular spine K Trapezius, transverse part Only the serratus posterior inferior is an intrinsic muscle of the back. The trapezius, latissimus dorsi, leva- tor scapulae, and rhomboideus muscles assist in move- ment of the shoulder or arm and are considered muscles of the upper limb. Fig. 2.1. Gilroy, MacPherson, Ross, Atlas of Anatomy, p. 22. Course of the Intercostal Nerves Copyright © 2008 by Thieme. All rights reserved. Illustrator: Markus Voll 39 Course of the Intercostal Nerves Coronal section. Anterior view. A Visceral pleura S Parietal pleura, diaphragmatic part D Diaphragm F Costodiaphragmatic recess G Endothoracic fascia H External intercostal J Costal groove K Intercostal vein, artery, and nerve Abnormal fluid collection in the pleural space (e.g., pleural effusion due to bronchial carcinoma) may necessitate the insertion of a chest tube. Generally, the optimal puncture site in a sitting patient is at the level of the 7th or 8th intercostal space on the posterior axillary line. The drain should always be introduced at the upper margin of a rib to avoid injuring the intercostal vein, artery, and nerve. Fig. 5.24. From Atlas of Anatomy, p. 59. Copyright ©2008 byThieme. Allrightsreserved. Illustrator: Markus Voll Right Lung 74 Right Lung Lateral and medial views. -

Gross Anatomy
www.BookOfLinks.com THE BIG PICTURE GROSS ANATOMY www.BookOfLinks.com Notice Medicine is an ever-changing science. As new research and clinical experience broaden our knowledge, changes in treatment and drug therapy are required. The authors and the publisher of this work have checked with sources believed to be reliable in their efforts to provide information that is complete and generally in accord with the standards accepted at the time of publication. However, in view of the possibility of human error or changes in medical sciences, neither the authors nor the publisher nor any other party who has been involved in the preparation or publication of this work warrants that the information contained herein is in every respect accurate or complete, and they disclaim all responsibility for any errors or omissions or for the results obtained from use of the information contained in this work. Readers are encouraged to confirm the infor- mation contained herein with other sources. For example and in particular, readers are advised to check the product information sheet included in the package of each drug they plan to administer to be certain that the information contained in this work is accurate and that changes have not been made in the recommended dose or in the contraindications for administration. This recommendation is of particular importance in connection with new or infrequently used drugs. www.BookOfLinks.com THE BIG PICTURE GROSS ANATOMY David A. Morton, PhD Associate Professor Anatomy Director Department of Neurobiology and Anatomy University of Utah School of Medicine Salt Lake City, Utah K. Bo Foreman, PhD, PT Assistant Professor Anatomy Director University of Utah College of Health Salt Lake City, Utah Kurt H. -

Diaphragm and Intercostal Muscles
Diaphragm and intercostal muscles Dr. Heba Kalbouneh Associate Professor of Anatomy and Histology Skeletal System Adult Human contains 206 Bones 2 parts: Axial skeleton (axis): Skull, Vertebral column, Thoracic cage Appendicular skeleton: Bones of upper limb Bones of lower limb Dr. Heba Kalbouneh Structure of Typical Vertebra Body Vertebral foramen Pedicle Transverse process Spinous process Lamina Dr. Heba Kalbouneh Superior articular process Intervertebral disc Dr. Heba Inferior articular process Dr. Heba Facet joints are between the superior articular process of one vertebra and the inferior articular process of the vertebra directly above it Inferior articular process Superior articular process Dr. Heba Kalbouneh Atypical Vertebrae Atlas (1st cervical vertebra) Axis (2nd cervical vertebra) Dr. Heba Atlas (1st cervical vertebra) Communicates: sup: skull (atlanto-occipital joint) inf: axis (atlanto-axial joint) Atlas (1st cervical vertebra) Characteristics: 1. no body 2. no spinous process 3. ant. & post. arches 4. 2 lateral masses 5. 2 transverse foramina Typical cervical vertebra Specific to the cervical vertebra is the transverse foramen (foramen transversarium). is an opening on each of the transverse processes which gives passage to the vertebral artery Thoracic Cage - Sternum (G, sternon= chest bone) -12 pairs of ribs & costal cartilages -12 thoracic vertebrae Manubrium Body Sternum: Flat bone 3 parts: Xiphoid process Dr. Heba Kalbouneh Dr. Heba Kalbouneh The external intercostal muscle forms the most superficial layer. Its fibers are directed downward and forward from the inferior border of the rib above to the superior border of the rib below The muscle extends forward to the costal cartilage where it is replaced by an aponeurosis, the anterior (external) intercostal membrane Dr. -
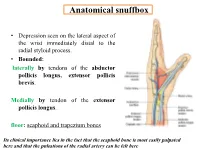
Anatomical Snuffbox
Anatomical snuffbox • Depression seen on the lateral aspect of the wrist immediately distal to the radial styloid process. • Bounded: laterally by tendons of the abductor pollicis longus, extensor pollicis brevis. Medially by tendon of the extensor pollicis longus. floor: scaphoid and trapezium bones Its clinical importance lies in the fact that the scaphoid bone is most easily palpated here and that the pulsations of the radial artery can be felt here Anatomical snuffbox Anatomical snuffbox • Contents: 2) Origin of the 1) The radial artery cephalic vein pass subcutaneously over the snuffbox. 3) Superficial branch of the radial nerve pass subcutaneously over the snuffbox. Blood supply of the hand Anastomoses occur between the radial and ulnar arteries via the superficial and deep palmar arches The Deep palmar arch is formed mainly by the radial artery while the superficial palmar arch is formed mainly by the ulnar artery 3-On entering the palm, it curves laterally behind (deep) the palmar 4-The arch is aponeurosis and in front completed on (superficial) of the long flexor the lateral side tendons forming by the the superficial palmar arch superficial branch of the radial artery. 2-Then it gives off its deep branch of which runs in front of the FR , and joins the radial artery to complete the deep palmar arch 1-Enters the hand anterior (superficial) to the Superficial flexor retinaculum palmar branch of radial artery through Guyon’s canal Radial artery 5-The superficial palmar arch gives off digital arteries from its convexity which pass to the fingers and supply them Superficial palmar arch Deep palmar branch of ulnar artery Superficial palmar branch of radial artery Ulnar artery Radial artery Radial Artery first dorsal interosseous muscle 1-From the floor of the anatomical snuff-box the radial artery leaves the dorsum of the hand by turning forward between the two heads of the first dorsal interosseous muscle. -

Brachium and Cubital Fossa
Anatomy Guy Dissection Sheet 1/15/2012 Brachium and Cubital Fossa Dr. Craig Goodmurphy Anatomy Guy Major Dissection Objectives – Anterior Compartment 1. Maintain the superficial veins but work the fascia of the brachium off the anterior compartment noting the intermuscular septae 2. Clean and identify the three muscle of the anterior arm and their attachments 3. Mobilize the contents of the brachial fascia as it extends from the axillary fascia to the elbow noting the median, ulnar and medial brachial and medial antebrachial cutaneous nerves 4. Follow the musculocutaneous nerve as it passes through the coracobrachialis and between the biceps and brachialis noting motor branches and the lateral antebrachial cutaneous nerve Major Dissection Objectives – Cubital Fossa & Posterior Compartment 6. Mobilize the cubital fossa veins and review the boundaries 7. Clean the biceps tendon and reflect the aponeurosis 8. Locate the contents of the fossa including the bifurcation of the brachial artery, median nerve and floor muscles 9. Have a partner elevate the arm to dissect posteriorly and remove the skin and fascia 10. Locate the three heads of the triceps and their attachments 11. Locate the profunda brachii artery and radial nerve at the triangular interval and between the brachialis and brachioradialis muscles Eastern Virginia Medical School 1 Anatomy Guy Dissection Sheet 1/15/2012 Brachium and Cubital Fossa Pearls & Problems Don’t 1. Cut the biceps muscle just mobilize it Do 2. Follow the cords and tubes from known to unknown as you clean them Do 3. Remove the duplicated deep veins but save the unpaired superficial veins Do 4. -

Questions on Human Anatomy
Standard Medical Text-books. ROBERTS’ PRACTICE OF MEDICINE. The Theory and Practice of Medicine. By Frederick T. Roberts, m.d. Third edi- tion. Octavo. Price, cloth, $6.00; leather, $7.00 Recommended at University of Pennsylvania. Long Island College Hospital, Yale and Harvard Colleges, Bishop’s College, Montreal; Uni- versity of Michigan, and over twenty other medical schools. MEIGS & PEPPER ON CHILDREN. A Practical Treatise on Diseases of Children. By J. Forsyth Meigs, m.d., and William Pepper, m.d. 7th edition. 8vo. Price, cloth, $6.00; leather, $7.00 Recommended at thirty-five of the principal medical colleges in the United States, including Bellevue Hospital, New York, University of Pennsylvania, and Long Island College Hospital. BIDDLE’S MATERIA MEDICA. Materia Medica, for the Use of Students and Physicians. By the late Prof. John B Biddle, m.d., Professor of Materia Medica in Jefferson Medical College, Phila- delphia. The Eighth edition. Octavo. Price, cloth, $4.00 Recommended in colleges in all parts of the UnitedStates. BYFORD ON WOMEN. The Diseases and Accidents Incident to Women. By Wm. H. Byford, m.d., Professor of Obstetrics and Diseases of Women and Children in the Chicago Medical College. Third edition, revised. 164 illus. Price, cloth, $5.00; leather, $6.00 “ Being particularly of use where questions of etiology and general treatment are concerned.”—American Journal of Obstetrics. CAZEAUX’S GREAT WORK ON OBSTETRICS. A practical Text-book on Midwifery. The most complete book now before the profession. Sixth edition, illus. Price, cloth, $6.00 ; leather, $7.00 Recommended at nearly fifty medical schools in the United States. -

Elbow Checklist
Workbook Musculoskeletal Ultrasound September 26, 2013 Shoulder Checklist Long biceps tendon Patient position: Facing the examiner Shoulder in slight medial rotation; elbow in flexion and supination Plane/ region: Transverse (axial): from a) intraarticular portion to b) myotendinous junction (at level of the pectoralis major tendon). What you will see: Long head of the biceps tendon Supraspinatus tendon Transverse humeral ligament Subscapularis tendon Lesser tuberosity Greater tuberosity Short head of the biceps Long head of the biceps (musculotendinous junction) Humeral shaft Pectoralis major tendon Plane/ region: Logitudinal (sagittal): What you will see: Long head of biceps; fibrillar structure Lesser tuberosity Long head of the biceps tendon Notes: Subscapularis muscle and tendon Patient position: Facing the examiner Shoulder in lateral rotation; elbow in flexion/ supination Plane/ region: longitudinal (axial): full vertical width of tendon. What you will see: Subscapularis muscle, tendon, and insertion Supraspinatus tendon Coracoid process Deltoid Greater tuberosity Lesser tuberosity Notes: Do passive medial/ lateral rotation while examining Plane/ region: Transverse (sagittal): What you will see: Lesser tuberosity Fascicles of subscapularis tendon Supraspinatus tendon Patient position: Lateral to examiner Shoulder in extension and medial rotation Hand on ipsilateral buttock Plane/ region: Longitudinal (oblique sagittal) Identify the intra-articular portion of biceps LH in the transverse plane; then -

H21/1 H21/2 H21/3
H21/1 (1013026) H21/2 (1013281) H21/1 H21/3 (1013282) H21/2 H21/3 (1013026/1013281/1013282) 2 Latin 1 Vertebra lumbalis [L V], processus articularis 51 Lig. supraspinale superior 52 Lig. sacroiliacum posterius 2 Vertebra lumbalis [L V], corpus vertebrae 53 Lig. sacrococcygeum laterale 3 Vertebra lumbalis [L V], processus costiformis; 54 Lig. sacrococcygeum posterius superficiale; processus costalis Lig. sacrococcygeum dorsale superficiale 4 Crista iliaca 55 Lig. sacrococcygeum posterius profundum; 5 Spina iliaca anterior superior Lig. sacrococcygeum dorsale profundum 6 Fossa iliaca 56 Foramen ischiadicum minus 7 Articulatio sacroiliaca 57 Canalis obturatorius 8 Spina iliaca anterior inferior 58 Arcus iliopectineus 9 Corpus ossis ilii 59 Lig. lacunare 10 Corpus ossis pubis 60 Lacuna vasorum 11 Fossa acetabuli 61 Lacuna musculorum 12 Spina ischiadica 62 Pars abdominalis aortae; Aorta abdominalis 13 Ramus ossis ischii 63 Vena cava inferior 14 Ramus superior ossis pubis 64 Truncus lumbosacralis 15 Ramus inferior ossis pubis 65 Ductus deferens 16 Discus interpubicus; Fibrocartilago interpubica 66 Arteria iliaca externa 17 Pecten ossis pubis 67 Vena iliaca externa 18 Foramen obturatum 68 M. cremaster 19 Foramina sacralia anteriora 69 Nn. scrotales anteriores 20 Promontorium 70 N. dorsalis penis 21 Ala ossis sacri 71 Glans penis 22 Articulatio lumbosacralis, discus intervertebralis 72 A. dorsalis penis® 23 Vertebra lumbalis [L V], processus articularis 73 V. dorsalis profunda penis inferior 74 Tunica vaginalis testis 24 Os sacrum; processus articularis superior 75 Epididymis 25 Ala ossis ilii 76 Plexus pampiniformis 26 Crista sacralis medialis 77 M. pyramidalis 27 Limbus acetabuli; Margo acetabuli 78 M. rectus abdominis 28 Foramen ischiadicum majus 79 Vesica urinaria 29 Tuber ischiadicum 80 M. -

Distal Radial Approach Through the Anatomical Snuff Box for Coronary Angiography and Percutaneous Coronary Intervention
Korean Circ J. 2018 Dec;48(12):1131-1134 https://doi.org/10.4070/kcj.2018.0293 pISSN 1738-5520·eISSN 1738-5555 Editorial Distal Radial Approach through the Anatomical Snuff Box for Coronary Angiography and Percutaneous Coronary Intervention Jae-Hyung Roh, MD, PhD, and Jae-Hwan Lee , MD, PhD Division of Cardiology, Department of Internal Medicine, Chungnam National University Hospital, Chungnam National University School of Medicine, Daejeon, Korea ► See the article “Feasibility of Coronary Angiography and Percutaneous Coronary Intervention via Left Snuffbox Approach” in volume 48 on page 1120. Received: Aug 27, 2018 The anatomical snuffbox, also known as the radial fossa, is a triangular-shaped depression Accepted: Sep 17, 2018 on the radial, dorsal aspect of the hand at the level of the carpal bones. It is clearly observed Figure 1 1)2) Correspondence to when the thumb is extended ( ). The bottom of the snuffbox is supported by carpal Jae-Hwan Lee, MD, PhD bones composed of the scaphoid and trapezium. The medial and lateral borders are bounded Division of Cardiology, Department of Internal by tendons of the extensor pollicis longus and the extensor pollicis brevis, respectively. The Medicine, Chungnam National University proximal border is formed by the styloid process of the radius. Within this narrow triangular Hospital, Chungnam National University space, various structures are located, including the distal radial artery (RA), a branch of the School of Medicine, 282, Munhwa-ro, Jung-gu, radial nerve, and the cephalic vein. Daejeon 35015, Korea. E-mail: [email protected] The anatomy of the hand arteries is illustrated in Figure 2. -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V.