Examinations of the Effectiveness of Mimetic Phenotypes of Female Great Mormom Butterfly Using Behavioral Experiments
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Butterflies-Of-Thailand-Checklist-2018
PAPILIONIDAE Parnassinae: Bhutanitis lidderdalii ocellatomaculata Great Bhutan ผเี สอื้ ภฐู าน Papilioninae: Troides helena cerberus Common Birdwing ผเี สอื้ ถงุ ทองป่ าสงู Troides aeacus aeacus Golden Birdwing ผเี สอื้ ถงุ ทองธรรมดา Troides aeacus malaiianus Troides amphrysus ruficollis Malayan Birdwing ผเี สอื้ ถงุ ทองปักษ์ใต ้ Troides cuneifera paeninsulae Mountain Birdwing ผเี สอื้ ถงุ ทองภเู ขา Atrophaneura sycorax egertoni Whitehead Batwing ผเี สอื้ คา้ งคาวหวั ขาว Atrophaneura varuna zaleucus Burmese Batwing ผเี สอื้ ปีกคา้ งคาวพมา่ Atrophaneura varuna varuna Malayan Batwing ผเี สอื้ ปีกคา้ งคาวมาเลย์ Atrophaneura varuna astorion Common Batwing ผเี สอื้ ปีกคา้ งคาวธรรมดา Atrophaneura aidoneus Striped Batwing ผเี สอื้ ปีกคา้ งคาวขา้ งแถบ Byasa dasarada barata Great Windmill ผเี สอื้ หางตมุ ้ ใหญ่ Byasa polyeuctes polyeuctes Common Windmill ผเี สอื้ หางตมุ ้ ธรรมดา Byasa crassipes Small Black Windmill ผเี สอื้ หางตมุ ้ เล็กด า Byasa adamsoni adamsoni Adamson's Rose ผเี สอื้ หางตมุ ้ อดัมสนั Byasa adamsoni takakoae Losaria coon doubledayi Common Clubtail ผเี สอื้ หางตมุ ้ หางกวิ่ Losaria neptunus neptunus Yellow-bodied Clubtail ผเี สอื้ หางตมุ ้ กน้ เหลอื ง Losaria neptunus manasukkiti Pachliopta aristolochiae goniopeltis Common Rose ผเี สอื้ หางตมุ ้ จดุ ชมพู Pachliopta aristolochiae asteris Papilio demoleus malayanus Lime Butterfly ผเี สอื้ หนอนมะนาว Papilio demolion demolion Banded Swallowtail ผเี สอื้ หางตงิ่ สะพายขาว Papilio noblei Noble's Helen ผเี สอื้ หางตงิ่ โนเบลิ้ Papilio castor mahadeva Siamese Raven ผเี สอื้ เชงิ ลายมหาเทพสยาม -

9 2013, No.1136
2013, No.1136 8 LAMPIRAN I PERATURAN MENTERI PERDAGANGAN REPUBLIK INDONESIA NOMOR 50/M-DAG/PER/9/2013 TENTANG KETENTUAN EKSPOR TUMBUHAN ALAM DAN SATWA LIAR YANG TIDAK DILINDUNGI UNDANG-UNDANG DAN TERMASUK DALAM DAFTAR CITES JENIS TUMBUHAN ALAM DAN SATWA LIAR YANG TIDAK DILINDUNGI UNDANG-UNDANG DAN TERMASUK DALAM DAFTAR CITES No. Pos Tarif/HS Uraian Barang Appendix I. Binatang Hidup Lainnya. - Binatang Menyusui (Mamalia) ex. 0106.11.00.00 Primata dari jenis : - Macaca fascicularis - Macaca nemestrina ex. 0106.19.00.00 Binatang menyusui lain-lain dari jenis: - Pteropus alecto - Pteropus vampyrus ex. 0106.20.00.00 Binatang melata (termasuk ular dan penyu) dari jenis: · Ular (Snakes) - Apodora papuana / Liasis olivaceus papuanus - Candoia aspera - Candoia carinata - Leiopython albertisi - Liasis fuscus - Liasis macklotti macklotti - Morelia amethistina - Morelia boeleni - Morelia spilota variegata - Naja sputatrix - Ophiophagus hannah - Ptyas mucosus - Python curtus - Python brongersmai - Python breitensteini - Python reticulates www.djpp.kemenkumham.go.id 9 2013, No.1136 No. Pos Tarif/HS Uraian Barang · Biawak (Monitors) - Varanus beccari - Varanus doreanus - Varanus dumerili - Varanus jobiensis - Varanus rudicollis - Varanus salvadori - Varanus salvator · Kura-Kura (Turtles) - Amyda cartilaginea - Calllagur borneoensis - Carettochelys insculpta - Chelodina mccordi - Cuora amboinensis - Heosemys spinosa - Indotestudo forsteni - Leucocephalon (Geoemyda) yuwonoi - Malayemys subtrijuga - Manouria emys - Notochelys platynota - Pelochelys bibroni -

PALATABILITAS KUPU-KUPU (LEPIDOPTERA: NYMPHALIDAE DAN PAPILIONIDAE) SEBAGAI PAKAN Tarsius Fuscus DI PENANGKARAN
Prosiding Seminar Nasional Biologi dan Pembelajarannya Universitas Negeri Medan, 12 Oktober 2018 ISSN 2656-1670 PALATABILITAS KUPU-KUPU (LEPIDOPTERA: NYMPHALIDAE DAN PAPILIONIDAE) SEBAGAI PAKAN Tarsius fuscus DI PENANGKARAN PALATABILITY OF BUTTERFLIES (LEPIDOPTERA: NYMPHALIDAE AND PAPILIONIDAE Tarsius fuscus IN CAPTIVITY Indra A.S.L.P. Putri1 1Balai Penelitian dan Pengembangan Lingkungan Hidup dan Kehutanan Makassar, e-mail: [email protected], Jalan Perintis Kemerdekaan Km 16.5 Telp +62411504049 dan +62411504058, Makassar ABSTRACT Tarsius fuscus is an endemic animal of Sulawesi. It is a nocturnal primate, consuming insects in its natural habitat. However, palatability of Nymphalidae and Papilionidae butterflies families as the feed of Tarsius fuscus is unknown. The butterflies are available abundantly around the Tarsius’ captivity in the natural habitat of Tarsius fuscus, therefore they are a potential natural feeding for Tarsius fuscus. This study aims to determine the palatability of Nymphalidae and Papilionidae butterflies families as the feed for Tarsius fuscus. This information is important to provide natural feeding for Tarsius fuscus, to avoid the dependence of Tarsius fuscus on grasshopper feed in captivity. The palatability treatment used was free choice feeding methods. The diet was given ad- libitum. The results showed that 12 species of butterflies from Nymphalidae family and 9 species of butterflies from the Papilionidae family were palatable for the feed of Tarsius fuscus Keywords : diet, Tarsius fuscus, Lepidoptera, captivity ABSTRAK Tarsius fuscus merupakan salah satu satwa endemik Sulawesi. Primata nocturnal ini diketahui mengkonsumsi berbagai spesies serangga di habitat alaminya, Namun demikian, palatabilitas Tarsius fuscus terhadap kupu-kupu dari familia Nymphalidae dan Papilionidae belum diketahui. Kupu-kupu ini diketahui tersedia cukup melimpah di sekitar lokasi kandang penangkaran yang di bangun di habitat alami Tarsius fuscus sehingga berpotensi tinggi sebagai pakan alami. -

An Inordinate Disdain for Beetles
An Inordinate Disdain for Beetles: Imagining the Insect in Colonial Aotearoa A Thesis submitted in partial fulfillment of the requirements for the Degree of Masters of Arts in English By Lillian Duval University of Canterbury August 2020 Table of Contents: TABLE OF CONTENTS: ................................................................................................................................. 2 TABLE OF FIGURES ..................................................................................................................................... 3 ACKNOWLEDGEMENT ................................................................................................................................ 6 ABSTRACT .................................................................................................................................................. 7 INTRODUCTION: INSECTOCENTRISM..................................................................................................................................... 8 LANGUAGE ........................................................................................................................................................... 11 ALICE AND THE GNAT IN CONTEXT ............................................................................................................................ 17 FOCUS OF THIS RESEARCH ....................................................................................................................................... 20 CHAPTER ONE: FRONTIER ENTOMOLOGY AND THE -

Checklist of Fish and Invertebrates Listed in the CITES Appendices
JOINTS NATURE \=^ CONSERVATION COMMITTEE Checklist of fish and mvertebrates Usted in the CITES appendices JNCC REPORT (SSN0963-«OStl JOINT NATURE CONSERVATION COMMITTEE Report distribution Report Number: No. 238 Contract Number/JNCC project number: F7 1-12-332 Date received: 9 June 1995 Report tide: Checklist of fish and invertebrates listed in the CITES appendices Contract tide: Revised Checklists of CITES species database Contractor: World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge, CB3 ODL Comments: A further fish and invertebrate edition in the Checklist series begun by NCC in 1979, revised and brought up to date with current CITES listings Restrictions: Distribution: JNCC report collection 2 copies Nature Conservancy Council for England, HQ, Library 1 copy Scottish Natural Heritage, HQ, Library 1 copy Countryside Council for Wales, HQ, Library 1 copy A T Smail, Copyright Libraries Agent, 100 Euston Road, London, NWl 2HQ 5 copies British Library, Legal Deposit Office, Boston Spa, Wetherby, West Yorkshire, LS23 7BQ 1 copy Chadwick-Healey Ltd, Cambridge Place, Cambridge, CB2 INR 1 copy BIOSIS UK, Garforth House, 54 Michlegate, York, YOl ILF 1 copy CITES Management and Scientific Authorities of EC Member States total 30 copies CITES Authorities, UK Dependencies total 13 copies CITES Secretariat 5 copies CITES Animals Committee chairman 1 copy European Commission DG Xl/D/2 1 copy World Conservation Monitoring Centre 20 copies TRAFFIC International 5 copies Animal Quarantine Station, Heathrow 1 copy Department of the Environment (GWD) 5 copies Foreign & Commonwealth Office (ESED) 1 copy HM Customs & Excise 3 copies M Bradley Taylor (ACPO) 1 copy ^\(\\ Joint Nature Conservation Committee Report No. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -

Diversity Pattern of Butterfly Communities (Lepidoptera
International Scholarly Research Network ISRN Zoology Volume 2011, Article ID 818545, 8 pages doi:10.5402/2011/818545 Research Article DiversityPatternofButterflyCommunities (Lepidoptera, Papilionoidae) in Different Habitat Types in a Tropical Rain Forest of Southern Vietnam Lien Van Vu1 and Con Quang Vu2 1 Department of Biology, Vietnam National Museum of Nature, 18 Hoang Quoc Viet, Nghia Do, Cau Giay, Hanoi, Vietnam 2 Department of Insect Ecology, Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet, Nghia Do, Cau Giay, Hanoi, Vietnam Correspondence should be addressed to Lien Van Vu, [email protected] Received 26 January 2011; Accepted 1 March 2011 Academic Editors: M. Griggio and V. Tilgar Copyright © 2011 L. V. Vu and C. Quang Vu. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Diversity of butterfly communities of a tropical rain forest of Bu Gia Map National Park in South Vietnam was studied in four different habitat types (the natural forest, the disturbed forest, the bamboo forest, and the stream sides in the forest) in December 2008 and April 2009. A total of 112 species with 1703 individuals of Papilionoidae (except Lycaenidae) were recorded. The proportion of rare species tends to decrease from the natural forest to the stream sides, while the proportion of common species tends to increase from the natural forest to the stream sides. The stream sides have the greatest individual number, while the disturbed forest contains the greatest species number. The bamboo forest has the least species and individual numbers. -

Barua Swallowtail Butterflies.Pmd
CATALOGUE ZOOS' PRINT JOURNAL 19(4): 1439-1441 PRESENT STATUS OF SWALLOWTAIL BUTTERFLIES IN GARBHANGA RESERVE FOREST, ASSAM, INDIA K.K. Barua, D. Kakati and J. Kalita Butterfly Research Centre, Department of Zoology, Guwahati University, Assam 781014, India. ABSTRACT METHODS Garbhanga Reserve forest in Assam has a rich diversity of Surveys were carried out at different spots within the study swallowtail butterflies with 29 species and subspecies belonging to eight genera. Habitat degradation caused by area by point and line transect methods (Barhaum et al., 1980, encroachment in fringe areas, illegal logging and stone 1981). The present survey was carried out from 2000 to 2002 quarrying is gradually posing a threat to the survival of these during the pre-monsoon, monsoon and the post-monsoon butterflies within the forest. seasons. KEYWORDS Butterflies were collected by chasing and netting as well as by Garbhanga, habitat, swallowtail butterflies, threat bait trapping as mentioned by the Zoological Survey of India (1990). Surveys were mostly carried out during the first half of The forests of Assam house a rich butterfly diversity. Evans the day. The collected specimens were preserved and identified (1932) described about 962 species and subspecies of butterflies following ZSI (1990), Evans (1932), Talbot (1939), Wynter-Blyth belonging to the five families from the Assam region alone. (1957), Mani (1986) and Haribal (1992). One specimen Swallowtails belonging to family Papilionidae are one of the representing each species and subspecies was preserved for most spectacular insects that have drawn the attention of future reference. A total of 70 individuals were collected and entomologists and naturalists. -

Six New Records of Butterflies from Lawachara National Park, Bangladesh
Tropical Natural History 16(2): 119-122, October 2016 2016 by Chulalongkorn University Short Note Six New Records of Butterflies from Lawachara National Park, Bangladesh AMIT KUMER NEOGI*, MD. SAMSUR RAHMAN, AFROZA SULTANA, ANIK CHANDRA MONDAL, TANVIR AHMED AND MD. NASIF SADAT Department of Zoology, Jagannath University, Dhaka-1100, BANGLADESH * Corresponding Author: Amit Kumer Neogi ([email protected]) Received: 19 December 2015; Accepted: 29 August 2016 Lawachara National Park (24°30΄- These are Burara oedipodea belesis Mabile, 24°32΄N, 91°37΄- 91°39΄E) is a mixed 1876; Tagiades menaka menaka Moore, tropical evergreen forest in Moulvibazar 1865; Suasa lisides lisides Hewitson, 1863; district, which lies under the north-eastern Arhopala fulla Hewitson, 1862; Flos diardi region of Bangladesh and considered as a diardi Hewitson, 1862; Atrophaneura hotspot of faunal diversity5. The lowland aidoneus Doubleday, 1845. This six new Sino-Himalayan butterflies are well record denotes previous sampling gaps in represented here and much of the tropical the area and suggest further study to fauna of Hong Kong and Guandong, compile a complete checklist to take southern China, and northern Indochina are conservation initiatives. also present in this area7. Butterfly research Burara oedipodea belesis (Mabile, on this kind of tropical area is still in the 1876; Branded Orange Awlet) was sighted ongoing process in Bangladesh. Lawachara in the coordinate of 24°20'11.83"N, National Park (LNP) originally supported an 91°48'58.48"E; from Moulvibazar (LNP) indigenous vegetation cover of mixed on 22 March 2014 at 6.00 am (GMT: tropical evergreen type1. Butterfly fauna of +06.00). -

Konservasi Alam Dlm Islam, 13-03
KONSERVASI ALAM DALAM ISLAM Konservasi Alam dalam Islam/Fachruddin M. Mangunjaya — Ed. 2 — Jakarta: Yayasan Obor Indonesia 2019 + hlm.; 14,5 x 21 cm ISBN: Judul: Konservasi Alam dalam Islam, Fachruddin M. Mangunjaya Copyrights © 2005 Diterbitkan pertama kali oleh Yayasan Obor Indonesia anggota IKAPI DKI Jakarta, atas bantuan Bank Dunia (The World Bank) dan Conservation International Indonesia Jakarta Edisi pertama: Mei 2005 Edisi kedua: YOI: Desain cover: Rahmatika Creative Design Yayasan Pustaka Obor Indonesia Jl. Plaju No. 10, Jakarta 10230, Indonesia T. + 62 (0)21 31926978, 31920114 F. + 62 (0)21 31924488 E-mail: yayasan_obor@cbn. net. id http://www.obor.or.id iv Mengajarkan ilmu pengetahuan dan menyebarkannya kepada yang belum mengerti tidak akan berkurang kecuali jika ilmu itu dirahasiakan (Umar bin Abdul Azis) Mengenang Kakeknda Allah yarham Al-Alamah Abdul Qadir Zailani al-Mentaya v This publication has been funded by The World Bank’s Faiths and Environment Initiative. The findings, interpretations, and conclusions expressed herein are those of the author(s) and do not necessarily reflect the views of the Executive Directors of the International Bank for Reconstruction and Development / The World Bank or the governments they represent. The World Bank does not guarantee the accuracy of the data included in this work. The boundaries, colors, denominations, and other information shown on any map in this work do not imply any judgment on the part of The World Bank concerning the legal status of any territory or the endorsement or acceptance of such boundaries. Penerbitan publikasi ini didukung oleh Inisiatif Keagamaan dan Lingkungan (Faiths and Environment Initiative) Bank Dunia. -

Of Dibang Valley, Mishmi Hills, Arunachal Pradesh, India
JoTT NOTE 4(12): 3137–3160 Butterflies (Lepidoptera) of Dibang is protected under Mehao Wildlife Valley, Mishmi Hills, Arunachal Sanctuary (93030’–95045’N & Pradesh, India 28005’–28015’E) (MWS) (Image 1). Monsoon Jyoti Gogoi The Dibang River originates in the western part of China and flows through this valley in a north- Centre for Biodiversity and Natural Resource Conservation, Department of Ecology and Environmental Science, Assam south direction. It is one of the main tributaries of the University, Silchar, Assam 788011, India Brahmaputra. Email: [email protected] The vegetation is characterized by tropical evergreen forests (up to 900m), subtropical and The Mishmi Hills (95049’–95059’ N & 28006’– temperate forests (900–1800 m), temperate broad leaf 28023’E) are located in the eastern region of Arunachal forest (1800–2800 m) and temperate conifer forest Pradesh. The altitude varies from 400–3568 m. The (2800–3500 m) (Champion & Seth 1968). hill range lies to the north of the Assam Valley. The major part of the Mishmi Hills is in the Dibang Methods Valley districts of Arunachal Pradesh. The Dibang Study Area: 1. Sally Lake (Image 2): The Lake is Valley districts have two divisions - Lower and Upper about 5km from Roing Town and lies at an altitude of Dibang Valley. The headquarters of the two districts about 400m. The lake is very close to the Deopani are Roing and Anini, respectively. Roing lies on River and is surrounded by hills. Small streams flow the plains near the Lohit River while Anini is at an through the bamboo forest into the open areas. -
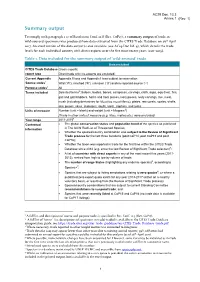
Summary Output
AC29 Doc. 13.3 Annex 1 Summary output To comply with paragraph 1 a) of Resolution Conf. 12.8 (Rev. CoP17), a summary output of trade in wild-sourced specimens was produced from data extracted from the CITES Trade Database on 26th April 2017. An excel version of the data output is also available (see AC29 Doc Inf. 4), which details the trade levels for each individual country with direct exports over the five most recent years (2011-2015). Table 1. Data included for the summary output of ‘wild-sourced’ trade Data included CITES Trade Database Gross exports; report type Direct trade only (re-exports are excluded) Current Appendix Appendix II taxa and Appendix I taxa subject to reservation Source codes1 Wild (‘W’), ranched (‘R’), unknown (‘U’) and no reported source (‘-’) Purpose codes1 All Terms included Selected terms2: baleen, bodies, bones, carapaces, carvings, cloth, eggs, egg (live), fins, gall and gall bladders, horns and horn pieces, ivory pieces, ivory carvings, live, meat, musk (including derivatives for Moschus moschiferus), plates, raw corals, scales, shells, skin pieces, skins, skeletons, skulls, teeth, trophies, and tusks. Units of measure Number (unit = blank) and weight (unit = kilogram3) [Trade in other units of measure (e.g. litres, metres etc.) were excluded] Year range 2011-20154 Contextual The global conservation status and population trend of the species as published information in The IUCN Red List of Threatened Species; Whether the species/country combination was subject to the Review of Significant Trade process for the last three iterations (post CoP14, post CoP15 and post CoP16); Whether the taxon was reported in trade for the first time within the CITES Trade Database since 2012 (e.g.