Rugose Corals Across the Devonian–Carboniferous Boundary in NW Turkey
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Masterarbeit
MASTERARBEIT Titel der Masterarbeit Late Cretaceous Volcaniclastic Rocks in the Pontides (NW Turkey) verfasst von Katharina Böhm BSc angestrebter akademischer Grad Master of Science (M.Sc.) Wien, 2015 Studienkennzahl lt. Studienblatt: A 066 815 Studienrichtung lt. Studienblatt: Erdwissenschaften Betreuerin / Betreuer: Ao. Univ.-Prof. Dr. Michael Wagreich Declaration ”I hereby declare that this master’s thesis was authored by myself independently, without use of other sources than indicated. I have explicitly cited all material which has been quoted either literally or by content from the used sources. Further this work was neither submitted in Austria nor abroad for any degree or examination.” Contents Declaration2 1. Introduction8 1.1. Project.................................. 9 1.1.1. Goals............................... 9 1.2. Geographical setting.......................... 10 1.3. Geological setting............................ 12 1.3.1. The Pontides........................... 14 1.3.2. The Pontides in the Cretaceous................ 18 1.3.3. Correlation with relative ages.................. 24 2. Nomenclature 28 3. Methods 29 3.1. ICP-ES and ICP-MS........................... 29 3.2. PXRD.................................. 29 3.3. Heavy mineral extraction........................ 30 4. Results 31 4.1. Mineralogy................................ 31 4.1.1. Powder X-Ray diffraction.................... 31 4.1.2. Thin sections.......................... 33 4.1.3. Mineral Extraction: Dating of minerals............. 34 4.2. Geochemistry.............................. 36 5. Interpretation of the geochemical results 37 5.1. Mobility of elements........................... 37 5.2. Alteration of minerals.......................... 39 5.3. Determining the rock type........................ 41 3 5.4. Discriminating volcanic series..................... 48 5.5. Revealing the tectonic setting...................... 52 5.6. Plotting geochemical element patterns................. 60 5.7. Summary of the geochemical classification.............. 62 6. -

Diapozitiv 1
Paleontologija vaje Aleksander Horvat in Luka Gale Spongia, Archaeocyatha, Cnidaria štud. l. 2008/09 PhylumPhylum PoriferaPorifera (spu(spužžve)ve) - mnogocelični organizmi, a sestojijo iz majhnega števila vrst celic, ki niso organizirane v tkiva, in nimajo živčevja, zato jih še nimamo za prave metazoje, temveč za parazoje; - parazoji niso predniki metazojev, temveč slepa evolucijska veja; - velika moč regeneracije, vendar celice ne morejo živeti samostojno; - sesilni bentos, filtratorji; -Največkrat imajo pokončno vrečasto telo z osrednjo votlino (paragaster), ki se navzven odpira z oskulom; zunanja površina je perforirana z drobnimi luknjicami (ostia), ki vodijo v dotočne kanale in kamre, ki so obdane s celicami ovratničarkami (hoanocite); iz kamer vodijo odtočni kanali do paragastra; - poleg ovratničark imajo nekatere še sploščene epitalialne celice (pinakocite), od katerih so nekatere perforirane (porocite) in lahko po potrebi zapirajo pore; skozi telo se prosto gibljejo amebocite, ki prenašajo hrano v druge dele spužve; -večina ima skelet: lahko je preprosta želatina, pri večini pa je iz roževinaste snovi (spongin) in/ali iz karbonatnih ali kremeničnih spikul; nekatere imajo poleg spikul še karbonatni skelet; arheociati so imeli le karbonaten skelet, brez spikul; - Pri recentnih spužvah ločimo tri stopnje organizacije: askon, sikon in levkon (ragon); - 1500 rec. Rodov, 80% morskih; - vir biogene kremenice, grebenotvorni organizmi (Cm – arheociati; Pz – stromatoporoidi; J – heksaktinelide…), bioerozija (Cliona); Glede na strukturo stene ločimo 2 poddebli: -Gelatinosa (zunanji epitalialni sloj leži preko želatinaste srednje plasti (mezenhim), v katerem so skleroblasti, ki izločajo spikule in kjer se gibljejo amebocite): -cl. Demospongea (kremenične spikule in/ali spongin): sklerospongije, hetetide, sfinktozoji, stromatoporoidi; - cl. Calcarea (Calcispongia) (karbonatne spikule): sfinktozoji, stromatoporoidi; - Nuda (nimajo niti zunanje plasti, niti mezenima): - cl. -

Multi-Phase Tectonic Movements and Their Controls on Coalbed Methane: a Case Study of No
energies Article Multi-Phase Tectonic Movements and Their Controls on Coalbed Methane: A Case Study of No. 9 Coal Seam from Eastern Yunnan, SW China Ming Li 1,2, Bo Jiang 1,2,* , Qi Miao 2,3, Geoff Wang 4, Zhenjiang You 4 and Fengjuan Lan 1,2 1 Key Laboratory of Coalbed Methane Resources and Reservoir Formation Process, Ministry of Education (China University of Mining and Technology), Xuzhou 221116, China; [email protected] (M.L.); [email protected] (F.L.) 2 School of Resources and Earth Science, China University of Mining and Technology, Xuzhou 221116, China; [email protected] 3 No. 198 Coalfield Geology Prospecting Team of Yunnan Province, Kunming 650208, China 4 School of Chemical Engineering, The University of Queensland, Brisbane, QLD 4072, Australia; [email protected] (G.W.); [email protected] (Z.Y.) * Correspondence: [email protected]; Tel.: +86-139-5135-9055 Received: 22 October 2020; Accepted: 16 November 2020; Published: 17 November 2020 Abstract: Multi-phase tectonic movements and complex geological structures limit the exploration and hotspot prediction of coalbed methane (CBM) in structurally complex areas. This scientific problem is still not fully understood, particularly in the Bumu region, Southwest China. The present paper analyses the occurrence characteristics and distribution of CBM based on the comprehensive analysis of CBM data. In combination with the analysis of the regional tectonics setting, geological structure features and tectonic evolution. The control action of multi-phase tectonic movements on CBM occurrence are further discussed. Results show that the Indosinian local deformation, Yanshanian intense deformation, and Himalayan secondary derived deformation formed the current tectonic framework of Enhong synclinorium. -

First Report of Myalinid Bivalves in the Lower Carboniferous of the Hakkari Basin, SE Turkey: Paleoecologic and Paleogeographic Implications
Turkish Journal of Earth Sciences Turkish J Earth Sci (2021) 30: 313-321 http://journals.tubitak.gov.tr/earth/ © TÜBİTAK Research Article doi:10.3906/yer-2010-20 First report of myalinid bivalves in the Lower Carboniferous of the Hakkari Basin, SE Turkey: paleoecologic and paleogeographic implications İzzet HOŞGÖR* Çalık Petrol (Oil and Gas Exploration and Production Company), Ankara, Turkey Received: 28.10.2020 Accepted/Published Online: 22.02.2021 Final Version: 17.05.2021 Abstract: Knowledge of the Late Paleozoic sedimentary history of the northern Gondwana shelf is advanced by new data from the Şort Dere section (Hakkari Basin). Early Carboniferous myalinid bivalves from the Middle East are reported here for the first time. The pre-Permian basement of the Hakkari Basin consists of a Lower Carboniferous sedimentary sequence including the Köprülü and Belek formations. The Köprülü Formation has yielded rich assemblages of vertebrates and invertebrates. Hitherto myalinid bivalves were unknown from this formation; this paper reports the discovery of small and poorly calcified myaliniform shells from two thin beds in the middle part of the Köprülü Formation of the Şort Dere section in the Zap Valley. The morphological characteristics of the specimens, especially the umbonal region with the numerous rounded wrinkles, suggest that it probably belongs to an undescribed species of Selenimyalina Newell, 1942. Based on this record, the state of the art of warm-water myalinid taxa in the Paleotethyan Realm is shortly discussed with a focus on the pteriomorph bivalve faunas within a global scenario for both paleobiogeography and patterns of Gondwana-Laurussia faunal exchange. The new occurrence is one of the most important pteriomorph bivalve records ever made in Southeast Turkey, helping to assist paleogeographic reconstructions of the Paleotethyan paleocontinents. -
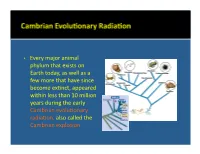
• Every Major Animal Phylum That Exists on Earth Today, As Well As A
• Every major animal phylum that exists on Earth today, as well as a few more that have since become ex:nct, appeared within less than 10 million years during the early Cambrian evolu:onary radiaon, also called the Cambrian explosion. • Phylum Pro:sta includes a diverse group of eukaryo:c microorganisms which are mostly unicellular. They are not necessarily primi:ve, although some are. Only some have a skeleton that can be preserved as a fossil. • Foraminifera and radiolaria are the most important examples. Coccolithophores, diatoms, and dinoflagellates are also examples. • Foraminifera are marine organisms that may be either planktonic, living in the water column where they float at various levels, or benthic, living on or within the seafloor sediment. • They form relavely large (0.1 mm to 5 cm) porous internal skeletons called tests through which project strands of living cytoplasm. The single- to mul:-chambered tests may be composed of organic maer, agglu:nated sand grains or sponge spicules, or calcium carbonate. • Benthic foraminifera • Planktonic foraminifera (Cambrian to Recent). (Jurassic to Recent). Oligocene Eocene • Radiolaria are planktonic marine organisms that tend to live in relavely cold water. • Their tests, oNen delicate and elaborate, are made of silica and presently accumulate on the floors of the parts of oceans that are deeper than those where foraminiferal tests are accumulang. • Radiolaria (Cambrian to • Radiolaria (Cambrian to Recent). Recent). Paleocene Radiolaria • Coccolithophores (Triassic to Recent) are extremely small marine planktonic photosynthe:c unicellular algae that are enclosed by plates of low-Mg calcite called coccoliths. Cretaceous White Cliffs of Dover, England • Diatoms (Jurassic to Recent) are small freshwater and marine planktonic photosynthe:c unicellular algae that are enclosed by a cell wall made of silica called a frustule. -

LCSH Section Z
Z (Computer program language) Zaan River Valley (Netherlands) Zabiello family (Not Subd Geog) [QA76.73.Z2] UF Zaan Valley (Netherlands) Here are entered works on families with the BT Programming languages (Electronic BT Valleys—Netherlands surnames Zabiello and Zabiełło. computers) Zaan Valley (Netherlands) When this heading is assigned to works on an individual family, the appropriate diacritical marks, if Z-49 (Video display terminal) USE Zaan River Valley (Netherlands) any, are included in the heading in the catalog record. USE Zenith Z-49 (Video display terminal) Zaar (African people) (May Subd Geog) UF Zabello family Z-80 (Microprocessor) [DT515.45.Z33] Zabirmawa (African people) USE Zilog Z-80 (Microprocessor) UF Gus (African people) USE Zarma (African people) Z-100 (Computer) Guus (African people) Zabirmawa language USE Zenith Z-100 (Computer) Saya (African people) USE Zarma language Z bosons Sayanci (African people) Zablan family (Not Subd Geog) [QC793.5.B62-QC793.5.B629] Sayara (African people) UF Sablan family UF Z particles Sayawa (African people) Zable family Z physics Seiyara (African people) USE Zabel family BT Bosons Seiyawa (African people) Zabludowski family (Not Subd Geog) Z Canyon (Wash.) Seya (African people) Zabon BT Canyons—Washington (State) Seyawa (African people) USE Pummelo Z-crank engines Sigdi (African people) Zaborski Park Krajobrazowy (Poland) USE Barrel engines Sigidi (African people) BT Parks—Poland Z-DNA Vigzar (African people) Zabrops (May Subd Geog) [QP624.5.Z33] Vikzar (African people) [QL537.A85] UF -

Prospects for Coal and Clean Coal Technologies in Turkey
Prospects for coal and clean coal technologies in Turkey Author: Dr Stephen Mills IEACCC Ref: CCC/239 ISBN: 978–92–9029–560–0 Copyright: © IEA Clean Coal Centre Published Date: July 2014 IEA Clean Coal Centre Park House 14 Northfields London SW18 1DD United Kingdom Telephone: +44(0)20 8877 6280 www.iea-coal.org IEA Clean Coal Centre – Prospects for coal and clean coal technologies in Turkey 2 Preface This report has been produced by IEA Clean Coal Centre and is based on a survey and analysis of published literature, and on information gathered in discussions with interested organisations and individuals. Their assistance is gratefully acknowledged. It should be understood that the views expressed in this report are our own, and are not necessarily shared by those who supplied the information, nor by our member countries. IEA Clean Coal Centre is an organisation set up under the auspices of the International Energy Agency (IEA) which was itself founded in 1974 by member countries of the Organisation for Economic Co-operation and Development (OECD). The purpose of the IEA is to explore means by which countries interested in minimising their dependence on imported oil can co-operate. In the field of Research, Development and Demonstration over fifty individual projects have been established in partnership between member countries of the IEA. IEA Clean Coal Centre began in 1975 and has contracting parties and sponsors from: Australia, Austria, Canada, China, the European Commission, Germany, India, Italy, Japan, New Zealand, Russia, South Africa, Thailand, the UK and the USA. The Service provides information and assessments on all aspects of coal from supply and transport, through markets and end-use technologies, to environmental issues and waste utilisation. -

Taxonomic Revision and Paleoecology of Middle Devonian (Eifelian) Fishes of the Onondaga, Columbus and Delaware Limestones of the Eastern United States
Graduate Theses, Dissertations, and Problem Reports 2002 Taxonomic revision and paleoecology of Middle Devonian (Eifelian) fishes of the Onondaga, Columbus and Delaware limestones of the eastern United States Robert Lewis Martin West Virginia University Follow this and additional works at: https://researchrepository.wvu.edu/etd Recommended Citation Martin, Robert Lewis, "Taxonomic revision and paleoecology of Middle Devonian (Eifelian) fishes of the Onondaga, Columbus and Delaware limestones of the eastern United States" (2002). Graduate Theses, Dissertations, and Problem Reports. 1678. https://researchrepository.wvu.edu/etd/1678 This Dissertation is protected by copyright and/or related rights. It has been brought to you by the The Research Repository @ WVU with permission from the rights-holder(s). You are free to use this Dissertation in any way that is permitted by the copyright and related rights legislation that applies to your use. For other uses you must obtain permission from the rights-holder(s) directly, unless additional rights are indicated by a Creative Commons license in the record and/ or on the work itself. This Dissertation has been accepted for inclusion in WVU Graduate Theses, Dissertations, and Problem Reports collection by an authorized administrator of The Research Repository @ WVU. For more information, please contact [email protected]. Taxonomic Revision and Paleoecology of Middle Devonian (Eifelian) Fishes of the Onondaga, Columbus and Delaware Limestones of the eastern United States. Robert L. Martin Dissertation submitted to the College of Arts and Sciences at West Virginia University in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Geology Thomas Kammer, Ph.D. -

Rugose Corals of the Donets Basin, Ukraine
TO L O N O G E I L C A A P I ' T A A T L E I I A Bollettino della Società Paleontologica Italiana, 59 (3), 2020, 205-224. Modena C N O A S S. P. I. Tournaisian (Carboniferous) rugose corals of the Donets Basin, Ukraine Viktor Ohar V. Ohar, Institute of Geology, Taras Shevchenko National University of Kyiv, 90 Vasylkivska str., 03022 Kyiv, Ukraine; [email protected] KEY WORDS - Rugosa, taxonomy, Dinantian. ABSTRACT - The results of the study of a rugose coral collection from the southern part of the Donets Basin are presented herein. Some species that were only mentioned in previous Donets Basin faunal lists are described and illustrated. The genera Aulokoninckophyllum Sando, 1976 and Merlewoodia Pickett, 1967 are reported for the first time from the Donets Basin. The stratigraphic distribution of the studied species is analysed. The uneven distribution of corals in the Tournaisian of the Donets Basin is explained by a complex combination of regional paleogeographic conditions and global events. The studied stratigraphic levels with corals are compared with the coral zones and third-order sequences of the Belgian Namur-Dinant Basin. INTRODUCTION in the foraminiferal evolutionary lineage Eoparastaffella (Devuyst et al., 2003), the assignment of the T/V boundary Tournaisian sedimentary rocks are exposed in the in the Donets Basin to the basal part of the Vb zone has southern part of the Donets Basin (Ukraine), where they been accepted. Thus, the Va zone is assigned now to are dominated by shallow-water platform limestone. It is the top of the Tournaisian. -

Exceptional New Fossil of Siphonophrentis Gigantea Christian Mccall
Exceptional new fossil of Siphonophrentis gigantea Christian McCall Abstract Rugose Corals, often referred to as Horn Coral, are an extinct order of stony corals in the phylum Cnidaria. They lived from the Ordivician period to the end of the Permian period, and can be found worldwide. A new fossil of Siphonophrentis gigantea, a species of Rugosa in the family Streptelasmatidae, has been recovered from the Devonian strata of the Lucas Formation. The fossil gives clues towards the paleobiology of Siphonophrentis, revealing it to have likely anchored itself to the sea bed in the ocean depths. Siphonophrentis gigantea likely had no relationship with Zooxanthellae, a kind of Dinoflagellate that gives modern extant coral their colour and allows them to photosynthesize. These single celled organisms appear to be absent in Siphonophrentis, and it instead received nutrients from a rich amount of biological debris that fell into its habitat. Further comparisons can be made between Siphonophrentis and the extant, cold-water coral Lophelia pertusa. Introduction Rugosa (Edwards et Haime, 1850), commonly known as “Horn Coral”, is an order of extinct marine animals that thrived from the Ordovician to the Permian period. They are called Horn Coral in reference to the concave depression (called a calice or calyx) on the tops of their fossils, which in life would have served as a living chamber for polyps. Rugosans ranged in size, with typical specimens usually preserving the calice and reaching a few centimetres in length. They can be identified by the pattern made by their septa, visible in the calice. Septa are the vertical plates inside the walls of the rugose coral, radiating from the centre (Sprung, 1999). -

Corals of Falls of the Ohio State Park
The Rugose Corals Tabulate Corals A Devonian coral garden Tabulae Rugose corals get their name because the exterior of Tabulate corals are always colonial. There are 212 species of corals many of their forms has a wrinkly appearance. They are Their name is derived from a flat shelf known from the fossil beds at the often called “horn corals” because their form may resemble (tabula) that separates individuals Falls of the Ohio. Not all occur in CORALS the horn of a cow or goat. In fact, the largest horn coral vertically in the colony (figure 6). The the coral beds; some may be found (Siphonophrentis elongata, figure 1) was referred to as a individual corallites that make up the in the slightly higher (younger) at “petrified buffalo horn” by colony are typically 0.5 - 8 mm wide. rock layers. Additional studies may Figure 11. Underside Falls of the Ohio settlers due to its shape. Figure 6. Despite the small individuals, these reveal more or fewer species. It takes of Alveolites coral Horn corals show a wide corals can be quite large. The most experience to identify corals with State Park variation in form, although common coral is a form called Favosites (Emmonsia). accuracy. The random orientation of many of the solitary external form is not a Emmonsia was given its own genus name, but some rugose corals make identification even more difficult. Alan Goldstein, Interpretive Naturalist distinguishing feature for specialists consider it to be a sub-genus of Favosites. These Look for evidence of Devonian hurricanes. Can you find identification. -

Heliophyllum a Study in Survival
Heliophyllum A Study in Survival Buffalo Geological Society Education Committee Bulletin – December 2019 ACKNOWLEDGEMENTS The Buffalo Geological Society thanks the members of our Education committee for their time and effort in producing what we hope to be the first of many informational bulletins on various facets of our hobby. Committee chairman Walter Drabek coordinated the project with the able assistance of several club members. Photography and photographic design were provided by Paul Leuchner whose nature photography is widely acclaimed in Western New York. Long time club member and amateur paleontologist Joseph Sullivan was responsible for technical review, enlisting the expertise of Dan Krisher of the Rochester Academy of Science and ensuring factual integrity. Mr. Krisher is a local coral expert who has worked with numerous organizations to produce fossil related publications and other educational events. Mark Castner of Canisius College provided editing and readability review as well as technical production support. Thin sections and acetate peels where produced at the Buffalo Museum of Science by Joseph Butch currently on the faculty at Buffalo State College and photographed by Michael Grenier of the Rochester Academy of Science. Specimens appearing in this pamphlet, that are not otherwise designated, are from the collections of various Club members. Finally, the Officers and Board need to be acknowledged for their support and critiquing of the final version. We dedicate this inaugural work to Bruce and Ruth Banick, Life Members of our organization and avid fossil collectors. The generous and ongoing commitment of their time and knowledge to our organization along with their extensive field work and the exhibition of their collections at numerous club events over the course of many years has inspired countless members to “keep on digging” and learning along the way.