Punctuated Equilibrium Comes of Age
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Paradox of Genius Recognition That, Although Nature Surely
BOOK REVIEWS recognition that, although nature surely embodies factual truth accessible to The paradox of genius human investigation (radical historians Stephen Jay Gould might even demur at this evident and necessary proposition), science does not and cannot 'progress' by rising above Darwin. By Adrian Desmond and James Moore. Michael Joseph: 1991. Pp. 808. social embeddedness on wings of a time £20. To be published in the United States by Warner Books later this year. less and international 'scientific method'. Science, like all human intellectual activ THE botanist J. D. Hooker, Charles kept throughout his life, especially as a ity, must proceed in a complex social, Darwin's closest friend and confidant, young man setting out to reform the political and psychological context: contributed a short preface to the great entire intellectual world. (These docu greatness must therefore be grasped as 1909 Cambridge Festschrift commemor ments had always been there, but were fruitful use, not as transcendence. ating both the fiftieth anniversary of the ignored and therefore unknown and in Darwin, so often depicted as a posi Origin of Species and the hundredth of visible.) I have lived ever since in the tivist hero, self-exiled at Downe in Kent Darwin's birth. Hooker, at the age of 92, penumbra of this industry as a practising amidst his plants and pigeons, emerges recalled his pleasure in Dar- win's trust and cited the volume of correspondence H.M.S. BEAGLE that he had received from MIDDLE SECTION FORE AND AFT Darwin alone: "upwards of a thousand pages of foolscap, each page containing, on an average, three hundred words." Darwin lived on that pre cious nineteenth-century crux of maximal information for historians - close enough to the present to permit pre r. -

The MBL Model and Stochastic Paleontology
216 Chapter seven ised exciting new avenues for research, that insights from biology and ecology could more profi tably be applied to paleontology, and that the future lay in assembling large databases as a foundation for analysis of broad-scale patterns of evolution over geological history. But in compar- ison to other expanding young disciplines—like theoretical ecology— paleobiology lacked a cohesive theoretical and methodological agenda. However, over the next ten years this would change dramatically. Chapter Seven One particular ecological/evolutionary issue emerged as the central unifying problem for paleobiology: the study and modeling of the his- “Towards a Nomothetic tory of diversity over time. This, in turn, motivated a methodological question: how reliable is the fossil record, and how can that reliability be Paleontology”: The MBL Model tested? These problems became the core of analytical paleobiology, and and Stochastic Paleontology represented a continuation and a consolidation of the themes we have examined thus far in the history of paleobiology. Ultimately, this focus led paleobiologists to groundbreaking quantitative studies of the inter- The Roots of Nomotheticism play of rates of origination and extinction of taxa through time, the role of background and mass extinctions in the history of life, the survivor- y the early 1970s, the paleobiology movement had begun to acquire ship of individual taxa, and the modeling of historical patterns of diver- Bconsiderable momentum. A number of paleobiologists began ac- sity. These questions became the central components of an emerging pa- tively building programs of paleobiological research and teaching at ma- leobiological theory of macroevolution, and by the mid 1980s formed the jor universities—Stephen Jay Gould at Harvard, Tom Schopf at the Uni- basis for paleobiologists’ claim to a seat at the “high table” of evolution- versity of Chicago, David Raup at the University of Rochester, James ary theory. -

Chordates (Phylum Chordata)
A short story Leathem Mehaffey, III, Fall 201993 The First Chordates (Phylum Chordata) • Chordates (our phylum) first appeared in the Cambrian, 525MYA. 94 Invertebrates, Chordates and Vertebrates • Invertebrates are all animals not chordates • Generally invertebrates, if they have hearts, have dorsal hearts; if they have a nervous system it is usually ventral. • All vertebrates are chordates, but not all chordates are vertebrates. • Chordates: • Dorsal notochord • Dorsal nerve chord • Ventral heart • Post-anal tail • Vertebrates: Amphioxus: archetypal chordate • Dorsal spinal column (articulated) and skeleton 95 Origin of the Chordates 96 Haikouichthys Myllokunmingia Note the rounded extension to Possibly the oldest the head bearing sensory vertebrate: showed gill organs bars and primitive vertebral elements Early and primitive agnathan vertebrates of the Early Cambrian (530MYA) Pikaia Note: these organisms were less Primitive chordate, than an inch long. similar to Amphioxus 97 The Cambrian/Ordovician Extinction • Somewhere around 488 million years ago something happened to cause a change in the fauna of the earth, heralding the beginning of the Ordovician Period. • Rather than one catastrophe, the late-Cambrian extinction seems to be a series of smaller extinction events. • Historically the change in fauna (mostly trilobites as the index species) was thought to be due to excessive warmth and low oxygen. • But some current findings point to an oxygen spike due perhaps to continental drift into the tropics, driving rapid speciation and consequent replacement of old with new organisms. 98 Welcome to the Ordovician YOU ARE HERE 99 The Ordovician Sea, 488 million years 100 ago The Ordovician Period lasted almost 45 million years, from 489 to 444 MYA. -

Punctuated Equilibrium Theory Variations in Punctuated Equilibrium and and the Diffusion of Innovation
An Introduction to Punctuated Equilibrium: A Model for Understanding Stability and Dramatic Change in Public Policies January 2018 This briefing note belongs to a series on the Tobacco policies can serve as an example to various models used in political science to illustrate this idea. Up until 1965, this policy had represent public policy development processes. changed very little, whereas in the late 1960s and Each of these briefing notes begins by describing early 1970s a radical change occurred in the analytical framework proposed by the given response to the actions of certain stakeholders, model. With this model in mind, we then set out to such as the US Surgeon General's 1964 examine questions that public health actors might publication of the now-famous report entitled ask about public policies. Our aim in these notes Smoking and Health. is not to further refine existing models; nor is it to advocate for the adoption of one model in To incorporate their insight into public policy particular. Our purpose is rather to suggest how analysis, Baumgartner and Jones sought to each of these models constitutes a useful reconcile in an integrated model the long periods interpretive lens that can guide reflection and of equilibrium, already well explained by the action leading to the production of healthy public incrementalist model, and the abrupt policies. punctuations of political systems. This became known as the punctuated equilibrium model. The punctuated equilibrium model aims to explain why public policies tend to be characterized by long periods of stability punctuated by short periods of radical change. -

V Sem Zool Punctuated Equilibrium
V Sem Zool Punctuated Equilibrium Gradualism and punctuated equilibrium are two ways in which the evolution of a species can occur. A species can evolve by only one of these, or by both. Scientists think that species with a shorter evolution evolved mostly by punctuated equilibrium, and those with a longer evolution evolved mostly by gradualism. Both phyletic gradualism and punctuated equilibrium are speciation theory and are valid models for understanding macroevolution. Both theories describe the rates of speciation. For Gradualism, changes in species is slow and gradual, occurring in small periodic changes in the gene pool, whereas for Punctuated Equilibrium, evolution occurs in spurts of relatively rapid change with long periods of non-change. The gradualism model depicts evolution as a slow steady process in which organisms change and develop slowly over time. In contrast, the punctuated equilibrium model depicts evolution as long periods of no evolutionary change followed by rapid periods of change. Both are models for describing successive evolutionary changes due to the mechanisms of evolution in a time frame. Punctuated equilibrium The punctuated equilibrium hypothesis states that speciation events occur rapidly in geological time - over hundreds of thousands to millions of years and that little change occurs in the time between speciation events. In other words, change only happens under certain conditions, and it happens rapidly. Instead of a slow, continuous movement, evolution tends to be characterized by long periods of virtual standstill or equilibrium punctuated by episodes of very fast development of new forms. It was proposed by Eldridge and Gould to explain the gaps in the fossil record - the fact that the fossil record does not show smooth evolutionary transitions. -

Multiple Molecular Evidences for a Living Mammalian Fossil
Multiple molecular evidences for a living mammalian fossil Dorothe´ e Huchon†‡, Pascale Chevret§¶, Ursula Jordanʈ, C. William Kilpatrick††, Vincent Ranwez§, Paulina D. Jenkins‡‡, Ju¨ rgen Brosiusʈ, and Ju¨ rgen Schmitz‡ʈ †Department of Zoology, George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv 69978, Israel; §Department of Paleontology, Phylogeny, and Paleobiology, Institut des Sciences de l’Evolution, cc064, Universite´Montpellier II, Place E. Bataillon, 34095 Montpellier Cedex 5, France; ʈInstitute of Experimental Pathology, University of Mu¨nster, D-48149 Mu¨nster, Germany; ††Department of Biology, University of Vermont, Burlington, VT 05405-0086; and ‡‡Department of Zoology, The Natural History Museum, London SW7 5BD, United Kingdom Edited by Francisco J. Ayala, University of California, Irvine, CA, and approved March 18, 2007 (received for review February 11, 2007) Laonastes aenigmamus is an enigmatic rodent first described in their classification as a diatomyid suggests that Laonastes is a 2005. Molecular and morphological data suggested that it is the living fossil and a ‘‘Lazarus taxon.’’ sole representative of a new mammalian family, the Laonastidae, The two research teams also disagreed on the taxonomic and a member of the Hystricognathi. However, the validity of this position of Laonastes. According to Jenkins et al. (2), Laonastes family is controversial because fossil-based phylogenetic analyses is either the most basal group of the hystricognaths (Fig. 2A)or suggest that Laonastes is a surviving member of the Diatomyidae, nested within the hystricognaths (Fig. 2B). According to Dawson a family considered to have been extinct for 11 million years. et al. (3), Laonastes and the other Diatomyidae are the sister According to these data, Laonastes and Diatomyidae are the sister clade of the family Ctenodactylidae (i.e., gundies), a family that clade of extant Ctenodactylidae (i.e., gundies) and do not belong does not belong to the Hystricognathi, but to which it is to the Hystricognathi. -

New York Ocean Action Plan 2016 – 2026
NEW YORK OCEAN ACTION PLAN 2016 – 2026 In collaboration with state and federal agencies, municipalities, tribal partners, academic institutions, non- profits, and ocean-based industry and tourism groups. Acknowledgments The preparation of the content within this document was developed by Debra Abercrombie and Karen Chytalo from the New York State Department of Environmental Conservation and in cooperation and coordination with staff from the New York State Department of State. Funding was provided by the New York State Environmental Protection Fund’s Ocean & Great Lakes Program. Other New York state agencies, federal agencies, estuary programs, the New York Ocean and Great Lakes Coalition, the Shinnecock Indian Nation and ocean-based industry and user groups provided numerous revisions to draft versions of this document which were invaluable. The New York Marine Sciences Consortium provided vital recommendations concerning data and research needs, as well as detailed revisions to earlier drafts. Thank you to all of the members of the public and who participated in the stakeholder focal groups and for also providing comments and revisions. For more information, please contact: Karen Chytalo New York State Department of Environmental Conservation [email protected] 631-444-0430 Cover Page Photo credits, Top row: E. Burke, SBU SoMAS, M. Gove; Bottom row: Wolcott Henry- 2005/Marine Photo Bank, Eleanor Partridge/Marine Photo Bank, Brandon Puckett/Marine Photo Bank. NEW YORK OCEAN ACTION PLAN | 2016 – 2026 i MESSAGE FROM COMMISSIONER AND SECRETARY The ocean and its significant resources have been at the heart of New York’s richness and economic vitality, since our founding in the 17th Century and continues today. -

The Tempo and Mode of Evolution Reconsidered Stephen Jay Gould
Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered Stephen Jay Gould; Niles Eldredge Paleobiology, Vol. 3, No. 2. (Spring, 1977), pp. 115-151. Stable URL: http://links.jstor.org/sici?sici=0094-8373%28197721%293%3A2%3C115%3APETTAM%3E2.0.CO%3B2-H Paleobiology is currently published by Paleontological Society. Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/journals/paleo.html. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academic journals and scholarly literature from around the world. The Archive is supported by libraries, scholarly societies, publishers, and foundations. It is an initiative of JSTOR, a not-for-profit organization with a mission to help the scholarly community take advantage of advances in technology. For more information regarding JSTOR, please contact [email protected]. http://www.jstor.org Sun Aug 19 19:30:53 2007 Paleobiology. -
Springer A++ Viewer
PublisherInfo PublisherName : BioMed Central PublisherLocation : London PublisherImprintName : BioMed Central Stephen Jay Gould dies ArticleInfo ArticleID : 4486 ArticleDOI : 10.1186/gb-spotlight-20020522-01 ArticleCitationID : spotlight-20020522-01 ArticleSequenceNumber : 152 ArticleCategory : Research news ArticleFirstPage : 1 ArticleLastPage : 3 RegistrationDate : 2002–5–22 ArticleHistory : OnlineDate : 2002–5–22 ArticleCopyright : BioMed Central Ltd2002 ArticleGrants : ArticleContext : 130593311 Hal Cohen Email: [email protected] PHILADELPHIA - Stephen Jay Gould, paleontologist, evolutionary biologist and popular author, died of abdominal mesothelioma Monday in New York City. He was 60. A provocative and controversial thinker, Gould was a fierce public defender of evolution. He became a figurehead for paleontology by making difficult concepts more digestible for the public in forums such as The New York Times and The New York Review of Books. While in graduate school, Gould and fellow student Niles Eldredge disputed the theories of evolution, which held that changes in organisms only occurred gradually, over eons. In their theory, known as punctuated equilibrium, evolution proceeded in bursts, followed by long periods of stasis. Thirty years later after their theory was first developed, the debate still rages. Among the many awards and honors bestowed upon Gould were membership in the National Academy of Sciences, the National Book Award, and the National Book Critics Circle Award. He also served as president of the American Association for the Advancement of Science. Gould wrote more than a dozen books, most recently publishing the 1,454-page The Structure of Evolutionary Theory (Harvard, 2001) and, this month, I Have Landed : The End of a Beginning in Natural History (Harmony Books, 2002). -

Punctuated Equilibrium Models in Organizational Decision Making 135
1 2 Punctuated Equilibrium Models 3 4 8 5 in Organizational Decision 6 7 Making 8 9 10 Scott E. Robinson 11 12 13 14 CONTENTS 15 16 8.1 Two Research Conundrums..................................................................................................134 17 8.1.1 Lindblom’s Theory of Administrative Incrementalism...........................................134 18 8.1.2 Wildavsky’s Theory of Budgetary Incrementalism.................................................135 19 8.1.3 The Diverse Meanings of Budgetary “Incrementalism”..........................................135 20 8.1.4 A Brief Aside on Paleontology.................................................................................136 21 8.2 Punctuated Equilibrium Theory—A Way Out of Both Conundrums.................................136 22 8.3 A Theoretical Model of Punctuated Equilibrium Theory....................................................137 23 8.4 Evidence of Punctuated Equilibria in Organizational Decision Making ............................139 24 8.4.1 Punctuated Equilibrium and the Federal Budget.....................................................139 25 8.4.2 Punctuated Equilibrium and Local Government Budgets.......................................140 26 8.4.3 Punctuated Equilibrium and the Federal Policy Process.........................................141 27 8.4.4 Punctuated Equilibrium and Organizational Bureaucratization ..............................142 28 8.4.5 Assessing the Evidence.............................................................................................143 -
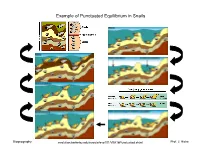
Example of Punctuated Equilibrium in Snails
Example of Punctuated Equilibrium in Snails Biogeography evolution.berkeley.edu/evosite/evo101/VIIA1bPunctuated.shtml1 Prof. J. Hicke Punctuated Equilibrium Lomolino et al. , 2006 Biogeography 2 Prof. J. Hicke Allopatric Speciation http://wps.pearsoncustom.com/wps/media/objects/3014/3087289/Web_Tutorials/18_A01.swf Biogeography 3 Prof. J. Hicke Allopatric Speciation: Vicariance Event Biogeography 4 Prof. J. Hicke Allopatric speciation, founder event Genes rare in original population are dominant in founding population Biogeography 5 Prof. J. Hicke Sympatric and Parapatric Speciation sympatric: extensive overlap parapatric: minimal overlap (partial geographic separation) Lomolino et al. , 2006 Biogeography 6 Prof. J. Hicke Parapatric Speciation No extrinsic barrier to gene flow, but… 1. restricted gene flow within population 2. varying selective pressures across the population range “Although continuously distributed, different flowering times have begun to reduce gene flow between metal-tolerant plants and metal-intolerant plants. “ evolution.berkeley.edu/evosite/evo101/VC1dParapatric.shtml Biogeography 7 Prof. J. Hicke Example of Sympatric Speciation • 200 years ago, flies only on hawthorns • then, introduction of domestic apple • females lay eggs on type of fruit they grew up on; males look for mates on type of fruit they grew up on • restricted gene flow • speciation http://evolution.berkeley.edu/evosite/evo101/VC1eSympatric.shtml Biogeography 8 Prof. J. Hicke Example of Sympatric Speciation Lomolino et al. , 2006 Biogeography 9 Prof. J. Hicke Adaptive Radiation often rapid speciation: Lake Victoria: 100s of new species in <12,000 years Lomolino et al. , 2006 Biogeography 10 Prof. J. Hicke Adaptive Radiation www.micro.utexas.edu/courses/levin/bio304/evolution/speciation.html Biogeography 11 Prof. -
Exaptation-A Missing Term in the Science of Form Author(S): Stephen Jay Gould and Elisabeth S
Paleontological Society Exaptation-A Missing Term in the Science of Form Author(s): Stephen Jay Gould and Elisabeth S. Vrba Reviewed work(s): Source: Paleobiology, Vol. 8, No. 1 (Winter, 1982), pp. 4-15 Published by: Paleontological Society Stable URL: http://www.jstor.org/stable/2400563 . Accessed: 27/08/2012 17:43 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Paleontological Society is collaborating with JSTOR to digitize, preserve and extend access to Paleobiology. http://www.jstor.org Paleobiology,8(1), 1982, pp. 4-15 Exaptation-a missing term in the science of form StephenJay Gould and Elisabeth S. Vrba* Abstract.-Adaptationhas been definedand recognizedby two differentcriteria: historical genesis (fea- turesbuilt by naturalselection for their present role) and currentutility (features now enhancingfitness no matterhow theyarose). Biologistshave oftenfailed to recognizethe potentialconfusion between these differentdefinitions because we have tendedto view naturalselection as so dominantamong evolutionary mechanismsthat historical process and currentproduct become one. Yet if manyfeatures of organisms are non-adapted,but available foruseful cooptation in descendants,then an importantconcept has no name in our lexicon (and unnamed ideas generallyremain unconsidered):features that now enhance fitnessbut were not built by naturalselection for their current role.