Thalictrum, Ranunculaceae) ⇑ Valerie L
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 5 Chapter 1 Introduction I. LIFE CYCLES and DIVERSITY of VASCULAR PLANTS the Subjects of This Thesis Are the Pteri
Chapter 1-15 Chapter 1 Introduction I. LIFE CYCLES AND DIVERSITY OF VASCULAR PLANTS The subjects of this thesis are the pteridophytes and seed plants that are conventionally classified as the vascular plants or tracheophytes. Vascular plants were traditionally defined by the possession of specialized conducting tissues, called phloem and xylem. Mosses are now believed to have inherited their conducting tissues from a common ancestor with the tracheophytes (Mishler & Churchill 1984) but are not considered in this thesis. Vascular plants can be divided into four groups with respect to life cycle. These groups are homosporous pteridophytes, heterosporous pteridophytes, gymnosperms and angiosperms. This is not intended to be a phylogenetic classification. There are about a quarter of a million species of vascular plant alive today. The vast majority are angiosperms and most of the remainder are homosporous pteridophytes. Heterosporous pteridophytes and gymnosperms contribute only a small number of species (Table 1.1). TABLE 1.1 Estimated number of extant species in each of the major groups of vascular plants (data from Parker 1982). Homosporous pteridophytes 12,000 species Heterosporous pteridophytes <1,000 species Gymnosperms <1,000 species Angiosperms >200,000 species A. Homosporous pteridophytes Homosporous pteridophytes produce a single type of spore. Spores are dispersed and develop into photosynthetic or mycoparasitic gametophytes. A gametophyte's gender is indeterminate at the time of spore dispersal, and a single gametophyte may produce eggs and/or sperm. Sperm are motile, and require free water to fertilize eggs. The young sporophyte is nourished by the maternal gametophyte during early development but later becomes Chapter 1-16 nutritionally independent. -

Floral MADS Box Genes and Homeotic Gender Dimorphism in Thalictrum Dioicum (Ranunculaceae) – a New Model for the Study of Dioecy
The Plant Journal (2005) 41, 755–766 doi: 10.1111/j.1365-313X.2005.02336.x Floral MADS box genes and homeotic gender dimorphism in Thalictrum dioicum (Ranunculaceae) – a new model for the study of dioecy Vero´ nica S. Di Stilio1,*,†, Elena M. Kramer1 and David A. Baum1,2 1Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138, USA, and 2Department of Botany, University of Wisconsin, Madison, WI 53706, USA Received 23 September 2004; revised 8 December 2004; accepted 14 December 2004. *For correspondence (fax þ206 616 2011; e-mail [email protected]). †Present address: Department of Biology, University of Washington, Seattle, WA 98195, USA. Summary In most dioecious angiosperm species, flowers are initially perfect but abort either stamens or carpels during their development, indicating that sex determination occurs after floral organ identity has been established. Dioecious members of the genus Thalictrum (meadow-rue), however, produce flowers that lack aborted organs. Examination of early flower development of T. dioicum confirms that flowers are male or female from inception, raising the possibility that genetic mechanisms working at or above the level of organ identity promote sex determination through a homeotic-like mechanism. In order to investigate this possibility, we identified homologs of the organ identity genes PISTILLATA (PI), APETALA3 (AP3) and AGAMOUS (AG) from T. dioicum and the hermaphroditic species T. thalictroides. A combination of early and late duplication events was uncovered in these gene lineages and expression analyses indicate that these events are generally associated with divergence in gene regulation. In light of these findings, we discuss the potential of T. -

Auxin Regulation Involved in Gynoecium Morphogenesis of Papaya Flowers
Zhou et al. Horticulture Research (2019) 6:119 Horticulture Research https://doi.org/10.1038/s41438-019-0205-8 www.nature.com/hortres ARTICLE Open Access Auxin regulation involved in gynoecium morphogenesis of papaya flowers Ping Zhou 1,2,MahparaFatima3,XinyiMa1,JuanLiu1 and Ray Ming 1,4 Abstract The morphogenesis of gynoecium is crucial for propagation and productivity of fruit crops. For trioecious papaya (Carica papaya), highly differentiated morphology of gynoecium in flowers of different sex types is controlled by gene networks and influenced by environmental factors, but the regulatory mechanism in gynoecium morphogenesis is unclear. Gynodioecious and dioecious papaya varieties were used for analysis of differentially expressed genes followed by experiments using auxin and an auxin transporter inhibitor. We first compared differential gene expression in functional and rudimentary gynoecium at early stage of their development and detected significant difference in phytohormone modulating and transduction processes, particularly auxin. Enhanced auxin signal transduction in rudimentary gynoecium was observed. To determine the role auxin plays in the papaya gynoecium, auxin transport inhibitor (N-1-Naphthylphthalamic acid, NPA) and synthetic auxin analogs with different concentrations gradient were sprayed to the trunk apex of male and female plants of dioecious papaya. Weakening of auxin transport by 10 mg/L NPA treatment resulted in female fertility restoration in male flowers, while female flowers did not show changes. NPA treatment with higher concentration (30 and 50 mg/L) caused deformed flowers in both male and female plants. We hypothesize that the occurrence of rudimentary gynoecium patterning might associate with auxin homeostasis alteration. Proper auxin concentration and auxin homeostasis might be crucial for functional gynoecium morphogenesis in papaya flowers. -

FLORA from FĂRĂGĂU AREA (MUREŞ COUNTY) AS POTENTIAL SOURCE of MEDICINAL PLANTS Silvia OROIAN1*, Mihaela SĂMĂRGHIŢAN2
ISSN: 2601 – 6141, ISSN-L: 2601 – 6141 Acta Biologica Marisiensis 2018, 1(1): 60-70 ORIGINAL PAPER FLORA FROM FĂRĂGĂU AREA (MUREŞ COUNTY) AS POTENTIAL SOURCE OF MEDICINAL PLANTS Silvia OROIAN1*, Mihaela SĂMĂRGHIŢAN2 1Department of Pharmaceutical Botany, University of Medicine and Pharmacy of Tîrgu Mureş, Romania 2Mureş County Museum, Department of Natural Sciences, Tîrgu Mureş, Romania *Correspondence: Silvia OROIAN [email protected] Received: 2 July 2018; Accepted: 9 July 2018; Published: 15 July 2018 Abstract The aim of this study was to identify a potential source of medicinal plant from Transylvanian Plain. Also, the paper provides information about the hayfields floral richness, a great scientific value for Romania and Europe. The study of the flora was carried out in several stages: 2005-2008, 2013, 2017-2018. In the studied area, 397 taxa were identified, distributed in 82 families with therapeutic potential, represented by 164 medical taxa, 37 of them being in the European Pharmacopoeia 8.5. The study reveals that most plants contain: volatile oils (13.41%), tannins (12.19%), flavonoids (9.75%), mucilages (8.53%) etc. This plants can be used in the treatment of various human disorders: disorders of the digestive system, respiratory system, skin disorders, muscular and skeletal systems, genitourinary system, in gynaecological disorders, cardiovascular, and central nervous sistem disorders. In the study plants protected by law at European and national level were identified: Echium maculatum, Cephalaria radiata, Crambe tataria, Narcissus poeticus ssp. radiiflorus, Salvia nutans, Iris aphylla, Orchis morio, Orchis tridentata, Adonis vernalis, Dictamnus albus, Hammarbya paludosa etc. Keywords: Fărăgău, medicinal plants, human disease, Mureş County 1. -

Alphabetical Lists of the Vascular Plant Families with Their Phylogenetic
Colligo 2 (1) : 3-10 BOTANIQUE Alphabetical lists of the vascular plant families with their phylogenetic classification numbers Listes alphabétiques des familles de plantes vasculaires avec leurs numéros de classement phylogénétique FRÉDÉRIC DANET* *Mairie de Lyon, Espaces verts, Jardin botanique, Herbier, 69205 Lyon cedex 01, France - [email protected] Citation : Danet F., 2019. Alphabetical lists of the vascular plant families with their phylogenetic classification numbers. Colligo, 2(1) : 3- 10. https://perma.cc/2WFD-A2A7 KEY-WORDS Angiosperms family arrangement Summary: This paper provides, for herbarium cura- Gymnosperms Classification tors, the alphabetical lists of the recognized families Pteridophytes APG system in pteridophytes, gymnosperms and angiosperms Ferns PPG system with their phylogenetic classification numbers. Lycophytes phylogeny Herbarium MOTS-CLÉS Angiospermes rangement des familles Résumé : Cet article produit, pour les conservateurs Gymnospermes Classification d’herbier, les listes alphabétiques des familles recon- Ptéridophytes système APG nues pour les ptéridophytes, les gymnospermes et Fougères système PPG les angiospermes avec leurs numéros de classement Lycophytes phylogénie phylogénétique. Herbier Introduction These alphabetical lists have been established for the systems of A.-L de Jussieu, A.-P. de Can- The organization of herbarium collections con- dolle, Bentham & Hooker, etc. that are still used sists in arranging the specimens logically to in the management of historical herbaria find and reclassify them easily in the appro- whose original classification is voluntarily pre- priate storage units. In the vascular plant col- served. lections, commonly used methods are systema- Recent classification systems based on molecu- tic classification, alphabetical classification, or lar phylogenies have developed, and herbaria combinations of both. -

Purple Meadow Rue, Thalictrum Dasycarpum
Purple Meadow Rue, Thalictrum dasycarpum Purple Meadow Rue is a delightful native perennial, 2-6 ft. high, that features blue- green lacy foliage (somewhat resembling columbine) and wiry-branched sprays of tiny, white flowers which bloom for 2-3 weeks in early summer (late May-July). The mass effect of the tiny flowers is quite showy! Emerging in loose, large clumps of soft, almost feathery sprays, the flowerheads can be a foot or more long, but have no true petals. Mostly dioecious (male/female flowers on separate plants), the male flowers are more fluffy with numerous yellow, thread-like stamens that droop out and become entangled. The female flowers are pollinated by wind and develop into spindle-shaped seeds that turn brown at maturity. A member of the buttercup family (Ranunculaceae), Purple Meadow Rue has an upright central stem that is unbranched below, but becomes branched above. The stems are usually purplish. The leaves are hairy beneath, alternate and compound: divided into numerous, three-lobed leaflets which become smaller as they ascend the stems. In the wild, Purple Meadow Rue is found in deciduous riparian woods, damp thickets and ravines, and moist prairies. Bees are sometimes attracted to the abundant pollen of the male flowers. However, this plant is not cross-pollinated by insects as the female flowers do not have nectar. There are a select few insect species that feed on this plant, including some aphids and caterpillars of certain moths. White-Tailed Deer browse the foliage sparingly. This species is currently being researched, because its seeds are one of the best sources of thalicarpine, a chemical with cancer-controlling properties. -

Etude Sur L'origine Et L'évolution Des Variations Florales Chez Delphinium L. (Ranunculaceae) À Travers La Morphologie, L'anatomie Et La Tératologie
Etude sur l'origine et l'évolution des variations florales chez Delphinium L. (Ranunculaceae) à travers la morphologie, l'anatomie et la tératologie : 2019SACLS126 : NNT Thèse de doctorat de l'Université Paris-Saclay préparée à l'Université Paris-Sud ED n°567 : Sciences du végétal : du gène à l'écosystème (SDV) Spécialité de doctorat : Biologie Thèse présentée et soutenue à Paris, le 29/05/2019, par Felipe Espinosa Moreno Composition du Jury : Bernard Riera Chargé de Recherche, CNRS (MECADEV) Rapporteur Julien Bachelier Professeur, Freie Universität Berlin (DCPS) Rapporteur Catherine Damerval Directrice de Recherche, CNRS (Génétique Quantitative et Evolution Le Moulon) Présidente Dario De Franceschi Maître de Conférences, Muséum national d'Histoire naturelle (CR2P) Examinateur Sophie Nadot Professeure, Université Paris-Sud (ESE) Directrice de thèse Florian Jabbour Maître de conférences, Muséum national d'Histoire naturelle (ISYEB) Invité Etude sur l'origine et l'évolution des variations florales chez Delphinium L. (Ranunculaceae) à travers la morphologie, l'anatomie et la tératologie Remerciements Ce manuscrit présente le travail de doctorat que j'ai réalisé entre les années 2016 et 2019 au sein de l'Ecole doctorale Sciences du végétale: du gène à l'écosystème, à l'Université Paris-Saclay Paris-Sud et au Muséum national d'Histoire naturelle de Paris. Même si sa réalisation a impliqué un investissement personnel énorme, celui-ci a eu tout son sens uniquement et grâce à l'encadrement, le soutien et l'accompagnement de nombreuses personnes que je remercie de la façon la plus sincère. Je remercie très spécialement Florian Jabbour et Sophie Nadot, mes directeurs de thèse. -

SIPARUNACEAE E MONIMIACEAE Siparuna Gêneros Pelos Reserva Na Representadas Estão Semelhantes
146 Siparunaceae e Monimiaceae As duas famílias foram recentemente monóicas ou dióicas. Os frutos são encobertos separadas. Monimiaceae sempre foi pelo receptáculo carnoso e quando maduros considerada heterogênea, e estudos atuais rompem-se irregularmente (com exceção de S. apontam para a segregação de certos gêneros glycycarpa, que é indeiscente), expondo drupas no nível de família, como o gênero Siparuna semelhantes a sementes, com arilo vermelho para Siparunaceae. São tratadas juntas neste ou alaranjado rico em lipídios. guia por serem vegetativamente semelhantes. As flores de Siparuna são polinizadas por Estão representadas na Reserva pelos gêneros moscas de hábito noturno, que visitam as flores Siparuna e Mollinedia. para acasalamento e oviposição. Monimiaceae é predominantemente pantropical com aproximadamente 194 espécies em ca. 22 gêneros, com a mesma distribuição de Siparunaceae na região neotropical. Na Amazônia ocorre somente o gênero Mollinedia, com uma espécie dióica, M. ovata registrada na Reserva. No campo a família é identificada através das mesmas características usadas para Siparunaceae. Mollinedia ovata tem folhas subcoriáceas, venação broquidódroma e poucas secundárias. As flores são como em Siparuna guianensis Siparunaceae, o receptáculo apresentando a Siparunaceae é neotropical e contém margem sinuosa. Os frutos são drupas sésseis aproximadamente 70 espécies em 2 gêneros ou estipitadas (semelhantes aos de Guatteria, (apenas um gênero mono-específico na Annonaceae) ou como em Siparunaceae, mas África Ocidental, Glossocalyx), distribuídas raramente com arilo alaranjado. desde o México e o Caribe até o Paraguai e Argentina. Somente Siparuna ocorre na Amazônia brasileira, estando representada na Reserva por nove espécies de hábito arbóreo ou arbustivo. O reconhecimento da família no campo é fácil: a lâmina foliar tem pontuações translúcidas e a planta tem um odor cítrico, agradável ou fétido (de onde vem o nome indígena caá-pitiú, ou planta com cheiro ruim), devido à grande concentração de óleos essenciais. -

Appendix 1. Species Summaries and Element Occurrence Records For
Appendix 1. Species summaries and element occurrence records for Campanula uniflora, Cymopterus evertii, Descurainia torulosa Helictotrichon hookeri, Papaver kluanense, and Thalictrum alpinum in the Carter Mountain Area of Critical Environmental Concern UYC»4ING NATURAL DIVERSITY DATABASE - The Nature Conservancy Key to Selected Fields in the Element Occurrence Database SNAME - The scientific name, in Latin, used in Wyoming (may be different in another state). SCOMNAME - The common name used in Wyoming. GRANK - The global rank assigned by TNC's network of Heritage Programs, based on world-wide distribution and threats. Ranks vary from G1, very rare or greatly threatened, through GS, common and secure. SRANK- The state rank assigned by each state Heritage Program, based on distribution within the state. Again, these ranks vary from S1, very rare or threatened, through SS, common and secure. These ranks may be different from state to state depending on the range of the taxon in each state. WYPLANT L!Sl-- WYNDD maintains a state lis~ where plants of special concern in the state appear on List 1, High Priority (rare, threatened, or endangered), List 2, Medium Priority, or List 3, Low Priority. PRECISION - The degree of refinement for an occurrence when it is mapped on a USGS quad; S=within seconds, M=within minutes, G=general (somewhere on the cited quad). COUNTYNAME - The county where the occurrence is located. QUADNAME - The name of the USGS 7.5 minute quad. MARG NUM indicates the dot on the map at the WYNDD office. LAT, LONG - Central latitude and longitude of the location if known from mapping procedures. -

Mountain Plants of Northeastern Utah
MOUNTAIN PLANTS OF NORTHEASTERN UTAH Original booklet and drawings by Berniece A. Andersen and Arthur H. Holmgren Revised May 1996 HG 506 FOREWORD In the original printing, the purpose of this manual was to serve as a guide for students, amateur botanists and anyone interested in the wildflowers of a rather limited geographic area. The intent was to depict and describe over 400 common, conspicuous or beautiful species. In this revision we have tried to maintain the intent and integrity of the original. Scientific names have been updated in accordance with changes in taxonomic thought since the time of the first printing. Some changes have been incorporated in order to make the manual more user-friendly for the beginner. The species are now organized primarily by floral color. We hope that these changes serve to enhance the enjoyment and usefulness of this long-popular manual. We would also like to thank Larry A. Rupp, Extension Horticulture Specialist, for critical review of the draft and for the cover photo. Linda Allen, Assistant Curator, Intermountain Herbarium Donna H. Falkenborg, Extension Editor Utah State University Extension is an affirmative action/equal employment opportunity employer and educational organization. We offer our programs to persons regardless of race, color, national origin, sex, religion, age or disability. Issued in furtherance of Cooperative Extension work, Acts of May 8 and June 30, 1914, in cooperation with the U.S. Department of Agriculture, Robert L. Gilliland, Vice-President and Director, Cooperative Extension -

2020 Inventory
2020 INVENTORY Wholesale Pricing at 30% Native Cultivar off Retail Available for Those in the Green Native Species Industry For A Complete Listing of Our Trees and Shrubs, Please Select and Purchase on Our On-Line Shop BLOOM PLANT NAME DESCRIPTION HT BLOOM EXPOSURE COST COLOR PERENNIALS blue green Actaea p. 'Misty Blue' 18" white june shade woodland native white sun/part Actaea racemosa see cimicifuga 6' July-Aug fragrant shade Agastache 'blue great long bloomer violet sun/part 36" July-Aug fortune' bottle brush blue shade bronze foliage violet Agastache 'bolero' 16'' July-Aug sun deer resistant pruple Agastache foeniculum anise hyssop 2-4' blue July-Aug sun Agastache 'Purple long blooming vibrant 3' July-Aug sun haze' native cultivar purple sterile pinwheel sun/part Allium 'Blue Eddy' 8-12" purple Sept rosettes shade lavender June- Allium cernuum nodding onion 1-2' sun pink August beautiful, fine 12- sun/part Amsonia 'Blue Ice' blue May-June foliage 15'' shade neeldle-like foliage sun/part Amsonia hubrichtii 3' blue May-June is golden in fall shade Amsonia 30- sun/part eastern blue star blue June tabernaemontana 36" shade white flowers, early sun/part Anemone canadensis 18'' white may spring shade 12- baby sun or Anemone 'Cinderella' vigorous clump fall 18'' pink shade 12- sun/part Anemone sylvestris spring beauty white April/May 18'' shade Antennaria pussy toes sun/part 6-12" white April/May plantaginifolia excellent ground shade shady woodland 30- shade/part Aquilegia ‘Nora Barlow’ mix April/May hummingbird plant 36'' shade shady rock -
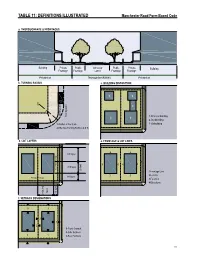
Manchester Road Redevelopment District: Form-Based Code
TaBle 11: deFiniTionS illuSTraTed manchester road Form-Based Code a. ThoroughFare & FronTageS Building Private Public Vehicular Public Private Building Frontage Frontage Lanes Frontage Frontage Private lot Thoroughfare (r.o.w.) Private lot b. Turning radiuS c. Building diSPoSiTion 3 3 2 2 1 Parking Lane Moving Lane 1- Principal Building 1 1 2- Backbuilding 1-Radius at the Curb 3- Outbuilding 2-Effective Turning Radius (± 8 ft) d. loT LAYERS e. FronTage & loT lineS 4 3rd layer 4 2 1 4 4 4 3 2nd layer Secondary Frontage 20 feet 1-Frontage Line 2-Lot Line 1st layer 3 3 Principal Frontage 3-Facades 1 1 4-Elevations layer 1st layer 2nd & 3rd & 2nd f. SeTBaCk deSignaTionS 3 3 2 1 2 1-Front Setback 2-Side Setback 1 1 3-Rear Setback 111 Manchester Road Form-Based Code ARTICLE 9. APPENDIX MATERIALS MBG Kemper Center PlantFinder About PlantFinder List of Gardens Visit Gardens Alphabetical List Common Names Search E-Mail Questions Menu Quick Links Home Page Your Plant Search Results Kemper Blog PlantFinder Please Note: The following plants all meet your search criteria. This list is not necessarily a list of recommended plants to grow, however. Please read about each PF Search Manchesterplant. Some may Road be invasive Form-Based in your area or may Code have undesirable characteristics such as above averageTab insect LEor disease 11: problems. NATIVE PLANT LIST Pests Plants of Merit Missouri Native Plant List provided by the Missouri Botanical Garden PlantFinder http://www.mobot.org/gardeninghelp/plantfinder Master Search Search limited to: Missouri Natives Search Tips Scientific Name Scientific Name Common NameCommon Name Height (ft.) ZoneZone GardeningHelp (ft.) Acer negundo box elder 30-50 2-10 Acer rubrum red maple 40-70 3-9 Acer saccharinum silver maple 50-80 3-9 Titles Acer saccharum sugar maple 40-80 3-8 Acer saccharum subsp.