On the Morphology of the Renal Tubules of Vertebrates
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Kidney, Renal Tubule – Dilation
Kidney, Renal Tubule – Dilation Figure Legend: Figure 1 Kidney, Renal tubule - Dilation in a male B6C3F1 mouse from a chronic study. Dilated tubules are noted as tracts running through the cortex and outer medulla. Figure 2 Kidney, Renal tubule - Dilation in a male F344/N rat from a chronic study. Tubule dilation is present throughout the outer stripe of the outer medulla, extending into the cortex. Figure 3 Kidney, Renal tubule - Dilation in a male B6C3F1 mouse from a chronic study. Slight tubule dilation is associated with degeneration and necrosis. Figure 4 Kidney, Renal tubule - Dilation in a male F344/N rat from a chronic study. Tubule dilation is associated with chronic progressive nephropathy. Comment: Renal tubule dilation may occur anywhere along the nephron or collecting duct system. It may occur in focal areas or as tracts running along the entire length of kidney sections (Figure 1). 1 Kidney, Renal Tubule – Dilation Renal tubule dilation may occur from xenobiotic administration, secondary mechanisms, or an unknown pathogenesis (see Kidney – Nephropathy, Obstructive (Figure 2). Dilation may result from direct toxic injury to the tubule epithelium interfering with absorption and secretion (Figure 3). It may also occur secondary to renal ischemia or from prolonged diuresis related to drug administration. Secondary mechanisms of tubule dilation may result from lower urinary tract obstruction, the deposition of tubule crystals, interstitial inflammation and/or fibrosis, and chronic progressive nephropathy (Figure 4). A few dilated tubules may be regarded as normal histologic variation. Recommendation: Renal tubule dilation should be diagnosed and given a severity grade. The location of tubule dilation should be included in the diagnosis as a site modifier. -

Anatomical and Morphological Study of the Kidneys of the Breeding Emu (Dromaius Novaehollandiae)
Turkish Journal of Zoology Turk J Zool (2016) 40: 314-319 http://journals.tubitak.gov.tr/zoology/ © TÜBİTAK Research Article doi:10.3906/zoo-1506-21 Anatomical and morphological study of the kidneys of the breeding emu (Dromaius novaehollandiae) 1, 2 3 2 3 Katarzyna MICHAŁEK *, Danuta SZCZERBIŃSKA , Marta GRABOWSKA , Danuta MAJEWSKA , Maria LASZCZYŃSKA 1 Department of Physiology, Cytobiology, and Proteomics, Faculty of Biotechnology and Animal Husbandry, West Pomeranian University of Technology in Szczecin, Szczecin, Poland 2 Department of Poultry and Ornamental Bird Breeding, West Pomeranian University of Technology in Szczecin, Szczecin, Poland 3 Department of Histology and Developmental Biology, Pomeranian Medical University, Szczecin, Poland Received: 15.06.2015 Accepted/Published Online: 05.01.2016 Final Version: 07.04.2016 Abstract: We analyzed 16 kidneys obtained from 15-year-old emus, Dromaius novaehollandiae. Emus were kept at an experimental farm in Poland. The results showed that each kidney was composed of 3 parts: cranial, medial, and caudal divisions. Histological results demonstrated that the kidneys consisted of 2 zones: the cortex and the medulla. The cortex made up the majority of the kidney, while the medulla formed only a small portion of the organ. Proximal and distal tubules and 2 types of glomeruli (looped and loopless) were localized in the cortex. Each of these glomeruli was characterized by tightly arranged mesangial cells. Proximal and distal tubules had a distinctive simple low cuboidal epithelium. The luminal surface of the proximal tubules had a brush border membrane, formed by numerous microvilli. The renal medulla of emu kidneys formed irregularly positioned characteristic cones of different sizes. -

Respiration and Elimination of Nitrogenous Wastes Forms and Functions of Plants and Animals
MODULE - 2 Respiration and Elimination of Nitrogenous Wastes Forms and Functions of Plants and animals 14 Notes RESPIRATION AND ELIMINATION OF NITROGENOUS WASTES Every living organism needs energy to perform various life activities, and the process of respiration fulfils this energy requirement. You have already learnt in the lesson on food and nutrition that animals take in high energy organic molecules in the form of food. During respiration, this food is broken down in the presence of oxygen and energy is released during respiration. Respiration also produces carbon dioxide, a toxic substance which is eliminated from the body. Thus, uptake of oxygen and removal of carbon dioxide is an essential requirement of all animals. At the same time numerous other toxic wastes such as ammonia, and urea are also produced in the tissues during various cellular activities. Such toxic wastes need to be removed from the body. In this lesson you will learn about removal of nitrogenous wastes and maintenance of water and salt balance in the body. OBJECTIVES After completing this lesson you will be able to : z define respiration, breathing, inspiration, expiration and vital capacity; z describe briefly the gaseous exchange in earthworm and cockroach; z describe the parts of respiratory system in the human body and mention their functions; z draw a labeled diagram of human respiratory system; z differentiate between breathing and respiration; and inspiration and expiration; z describe the mechanism of breathing and its regulation; z describe the exchange -

Morphology of the Kidney in a Nectarivorous Bird, the Anna's Hummingbird Calypte Anna
J. Zool., Lond. (1998) 244, 175±184 # 1998 The Zoological Society of London Printed in the United Kingdom Morphology of the kidney in a nectarivorous bird, the Anna's hummingbird Calypte anna G. Casotti1*, C. A. Beuchat2 and E. J. Braun3 1 Department of Biology, West Chester University, West Chester, PA, 19383, U.S.A. 2 Department of Biology, San Diego State University, San Diego, CA, 92182, U.S.A. 3 Department of Physiology, Arizona Health Sciences Center, University of Arizona, P.O. Box 245051, Tucson, AZ, 85724±5051, U.S.A. (Accepted 21 April 1997) Abstract The kidneys of Anna's hummingbird (Calypte anna) differ in several signi®cant ways from those of other birds that have been examined. The kidneys of this nectarivore contain very little medullary tissue; 90% of the total volume of the kidneys is cortical tissue, with medulla accounting for only an additional 2%. More than 99% of the nephrons are the so-called `reptilian type', which lack the loop of Henle. The few looped (`mammalian type') nephrons are incorporated into only a few medullary cones per kidney. The loopless nephrons are similar to those of other birds. However, the looped nephrons differ in that they lack the thin descending limb of the loop of Henle, which is found in other birds and is thought to play an important role in the countercurrent multiplier system in the avian kidney. Instead, the cells of the nephron segment following the pars recta of the proximal tubule resemble those of the thick ascending limb, with the large populations of mitochondria that are typical of transporting epithelia and no reduction in cell height. -
Evaluating and Treating the Kidneys
16_Nephrology.qxd 8/23/2005 10:41 AM Page 451 CHAPTER 16 Evaluating and Treating the Kidneys M. SCOTT ECHOLS, DVM, D ipl ABVP-A vian Renal diseases and their various classifications are well documented in the avian literature. The precise patho- genesis of avian renal disease, however, is not nearly as well described as it is in mammals. Renal disease has been shown to be fairly common in avian species. In studied poultry, as much as 29.6% of all disease condi- tions had abnormal pathology associated with or attrib- utable to renal disorders.20,214,251 Amyloidosis, urate nephrosis and gout were the most common diseases associated with mortality in a 4-year retrospective study conducted at a waterfowl park in Ontario, Canada.210 Thirty-seven percent of all avian cases presenting with renal tissue for histopathological examination, included over a 15-month period at the Schubot Exotic Bird Health Center, had one or more histologically identified kidney lesions.187 Nine of 75 pheasants (Phasianus colchicus) died with nephritis, one or both ureters impacted, and visceral gout in another comprehensive study on avian mortality.186 These case reports and retro- spective studies support the conclusion that renal dis- eases are relatively common and are clinically significant in multiple avian species. When compared to mammalian counterparts, the avian urogenital system has many structural and functional differences, which have been described previously.77,90,118,181,187,227 Differences including gross anatomy, renal portal blood flow and protein waste elimination should be considered when reviewing this chapter, as findings obtained from mammalian studies may not necessarily be applicable to birds. -

Seminiferous Tubules to Epididymis in the Mouse: a Histological and Quantitative Study
TRANSPORT OF SPERMATOZOA FROM SEMINIFEROUS TUBULES TO EPIDIDYMIS IN THE MOUSE: A HISTOLOGICAL AND QUANTITATIVE STUDY BRUCE M. BARACK Department of Anatomy, Washington University School of Medicine, St. Louis, Missouri, U.S.A. (Received 1st May 1967) Summary. The histology and structure of the seminiferous tubules, the tubuli recti, the rete testis and the ductuli efferentes of the adult albino mouse were studied in conjunction with the effect of unilateral ligation of the ductuli efferentes on the testis of the operated side under various experimental conditions. Results indicate that the amount of fluid produced within the testis is sufficient to flush the tubular system at least once a day and therefore eight times/spermatogenic cycle and that this fluid passage is the major factor in the transport of spermatozoa from the seminiferous tubules to the ductuli efferentes. The possibility of an interaction between the Sertoli cell and associated spermatids in early spermiogenesis resulting in secretion of fluid by the Sertoli cell is discussed. INTRODUCTION The mechanism of transport of immotile spermatozoa from the seminiferous tubules to the epididymis is still in question. The consequences of unilateral ligation of the ductuli efferentes or the head of the epididymis on the testis of the operated side have been described by Van Wagenen (1924, 1925), White (1933), Harrison (1953) and Harrison & Macmillan (1954) in the rat; Ladman & Young (1958) in the guinea-pig; Baillie (1962) in the mouse and Gaddum & Glover (1965) in the rabbit. These investigators found that following ligation there occurs a generalized dilatation of the seminiferous tubules accompanied by a varying degree of shedding and/or degeneration of the seminiferous epithelium. -
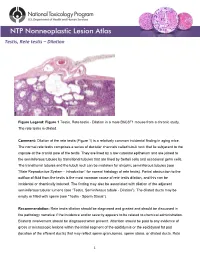
Testis, Rete Testis – Dilation
Testis, Rete testis – Dilation Figure Legend: Figure 1 Testis, Rete testis - Dilation in a male B6C3F1 mouse from a chronic study. The rete testis is dilated. Comment: Dilation of the rete testis (Figure 1) is a relatively common incidental finding in aging mice. The normal rete testis comprises a series of ductular channels called tubuli recti that lie subjacent to the capsule at the cranial pole of the testis. They are lined by a low cuboidal epithelium and are joined to the seminiferous tubules by transitional tubules that are lined by Sertoli cells and occasional germ cells. The transitional tubules and the tubuli recti can be mistaken for atrophic seminiferous tubules (see “Male Reproductive System - Introduction” for normal histology of rete testis). Partial obstruction to the outflow of fluid from the testis is the most common cause of rete testis dilation, and this can be incidental or chemically induced. The finding may also be associated with dilation of the adjacent seminiferous tubular lumens (see “Testis, Seminiferous tubule - Dilation”). The dilated ducts may be empty or filled with sperm (see “Testis - Sperm Stasis”). Recommendation: Rete testis dilation should be diagnosed and graded and should be discussed in the pathology narrative if the incidence and/or severity appears to be related to chemical administration. Bilateral involvement should be diagnosed when present. Attention should be paid to any evidence of gross or microscopic lesions within the initial segment of the epididymis or the epididymal fat pad (location of the efferent ducts) that may reflect sperm granulomas, sperm stasis, or dilated ducts. Rete 1 Testis, Rete testis – Dilation testis dilation may be associated with seminiferous tubular dilation and/or germinal epithelial atrophy, since these may be a consequence of obstruction of fluid outflow. -

The “Road Map”
PRACTICAL ROADMAP MALE REPRODUCTIVE SYSTEM DR N GRAVETT THE TESTIS • Slide 7 Stain: Iron Haematoxylin NOTE: Iron haematoxylin, a blue-black stain demonstrates the chromosomes in the dividing cells of the testis THE TESTIS Connective Tissue Septum These incomplete septae Tunica Albuginea divide the testis into lobes Seminiferous Tubule Interstitial Tissue Loose connective tissue between the seminiferous tubules THE TESTIS Tunica Tunica Albuginea Vasculosa BV Seminiferous Tubule Leydig Cells Blood Vessel (BV) Interstitial Seminiferous Tubule Tissue LEYDIG CELLS Interstitial Tissue BV Seminiferous Tubule NOTE: Leydig cells are endocrine glands and as such are usually located close to blood vessels. These cells are located outside the seminiferous tubules within the loose connective tissue stroma. SEMINIFEROUS TUBULE • Seminiferous Epithelium – Complex Stratified Epithelium consisting of 2 basic cell populations: 1. Sertoli Cells 2. Cells of the Spermatogenic Series: • Spermatogonia • Primary Spermatocyte • Secondary Spermatocyte (Transitory phase: not seen in histological section) • Early Spermatid • Late Spematid SEMINIFEROUS TUBULE Myoid Cell Sertoli Cells Primary Spermatocyte Spermato- gonium Spermato- gonium Lumen Early Spermatids Late Spematids Leydig Cell Spermato- gonium TESTIS AND EPIDIDYMIS • Slide 11 Stain: H&E NOTE: This slide is for ANAT 2020 only Pathway of sperm from point of production to exterior: Seminiferous Tubule Tubuli recti Rete Testes Efferent Ductules Epididymis Vas Deferens Ejaculatory Duct Prostatic Urethra -
Animal/Dairy Science 434� Learning Objectives
Animal/Dairy Science 434 ! Learning Objectives Male Reproductive Tract Anatomy! • To be able to describe the male reproductive system in the bull. – Structure – Function Testis! Epididymis! Factory! Finishing School! Bull Reproductive Tract! • 1-25 X 109 sperm/day! • Fluid Absorption! • “Plant” must be air • 8-25 X 109 sperm! conditioned! – membrane changes! – nuclear & flagellar stabilization! – motility, fertility! Penis! – cytoplasmic droplet translocation! Delivery System! • Erection! • Protrusion! • Ejaculation! Tail of Epididymis! Warehouse and Delivery! • Storage 10-50 X 109 sperm! • Sperm for 5-10 ejaculates! • Smooth muscle contractions Accessory Sex Glands! upon sexual stimulation! Alterations & Packaging! • Metabolic substrates! • Surface coatings! • Transport for sperm! Reproductive Reproductive Organs of the Organs of the Bull! Bull! Testis! Primary Sex Organ Bull Tract! • Thermosensor! • Radiator! Bull Testis! • Protective sac! Scrotum! Bull Tract! Scrotal Cross Section! Longitudinal Section Cross Section Structure of the Testis! Scrotal Longitudinal" Seminiferous Tubule Section ! Tunica Albuginea Rete Testis (within the mediastinum) Structure of Mediastinum! Rete Testis! the Testis! Seminiferous! Tubule! Seminiferous Tubule Tunica Albuginea Rete Testis (within the mediastinum) The Testis! Structure of Spermatic Cord Structure of Spermatic Cord the Testis! the Testis! Secondary Sex Organs Vas Deferens Vas Deferens Caput Epididymis Caput Epididymis • Vasectomy • Vasectomy Vas Efferentia Vas Efferentia Seminiferous Tubule 6-12 -

Histological Examination of Testicular Cell Development and Apoptosis in the Ostrich Chick
L. WEI, K. M. PENG, H. LIU, H. SONG, Y. WANG, L. TANG Turk. J. Vet. Anim. Sci. 2011; 35(1): 7-14 © TÜBİTAK Research Article doi:10.3906/vet-0806-2 Histological examination of testicular cell development and apoptosis in the ostrich chick Lan WEI1,2, Ke-Mei PENG1,*, Huazhen LIU1, Hui SONG1, Yan WANG1, Lia TANG1 1Department of Anatomy, Histology and Embryology, College of Animal Science and Veterinary Medicine, Huazhong Agricultural University, Wuhan 430070 - CHINA 2College of Animal Science and Technology, Henan University of Science and Technology, Luoyang 471003 - CHINA Received: 06.06.2008 Abstract: Th e aim of this study was to observe the microstructure and ultrastructure of the testis, demonstrate testicular cell development and apoptosis, and elucidate regularity of development and apoptosis in ostrich chick testes. By employing light microscopy 3 obvious development characteristics were detected with ostrich age increasing: fi rst, many primordial germ cells and a few spermatogonia were found while seminiferous tubule integrity was not evident in testes of 1-day-old ostrich chicks nor were primordial germ cells. In the 30-day-old ostrich chicks spermatogonia were completely diff erentiated, but very few primary spermatocytes were observed in testes at 45 days of age. Second, the quantity of mitochondria in spermatogonia and lipid droplet in Leydig cells increased gradually with the increasing age of astrish. Th ird, testicular cell apoptosis was observed and the number of apoptotic testicular cells showed a peak in the testis of 45-day-old ostrich chicks (P < 0.05), as testicular cells were prone to apoptosis at that age. -

Renal Medullary Tubule Organization Causes Immaturity of Urine-Concentrating Ability in Neonates
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Kidney International, Vol. 60 (2001), pp. 680–693 “Avian-type” renal medullary tubule organization causes immaturity of urine-concentrating ability in neonates WEN LIU,1 TETSUJI MORIMOTO,1 YOSHIAKI KONDO,KAZUIE IINUMA,SHINICHI UCHIDA, and MASASHI IMAI Department of Pediatrics, Tohoku University School of Medicine, Sendai, Miyagi; Second Department of Internal Medicine, Tokyo Medical and Dental University, Tokyo; and Department of Pharmacology, Jichi Medical College, Kawachi, Tochigi, Japan “Avian-type” renal medullary tubule organization causes im- cating the absence of urea-dependent volume flux in the neona- maturity of urine-concentrating ability in neonates. tal IMCD. These transport characteristics in each neonatal Background. While neonatal kidneys are not powerful in tubule are similar to those in quail kidneys. Identification of concentrating urine, they already dilute urine as efficiently mRNAs and immunofluorescent studies for specific transport- as adult kidneys. To elucidate the basis for this paradoxical ers, including rAQP-1, rCCC2, rCLC-K1, rENaC  subunit, immaturity in urine-concentrating ability, we investigated the rAQP-2, and rUT-A1, supported these findings. function of Henle’s loop and collecting ducts (IMCDs) in the Conclusion. We hypothesize that the renal medullary tubule inner medulla of neonatal rat kidneys. organization of neonatal rats shares a tremendous similarity Methods. Analyses of individual renal tubules in the inner with avian renal medulla. The qualitative changes in the organi- medulla of neonatal and adult rat kidneys were performed zation of medullary tubules may be primarily responsible for by measuring mRNA expression of membrane transporters, the immature urine-concentrating ability in mammalian neo- transepithelial voltages, and isotopic water and ion fluxes. -

RESPIRATORY SYSTEM Parts of Respiratory System
Chapter 2 Respiratory System RESPIRATORY SYSTEM The respiratory system plays role in exchange of oxygen and carbon dioxide between the atmospheric air, blood and tissue cells. Metabolic reactions in human body use oxygen (O2) to produce energy from nutrients in form of ATP. At the same time, these reactions release carbon dioxide (CO2) which must be eliminated quickly to prevent acidity due to CO2. The respiratory and cardiovascular systems cooperate to supply O2 and eliminate CO2. The respiratory system provides for gas exchange i.e. intake of O2 and elimination of CO2. The cardiovascular system transports these gases through blood between the lungs and body cells. The respiratory system also participates in regulating blood pH, contains receptors for the sense of smell, filters inspired air, produces sounds, and rids the body of some water and heat in exhaled air. Parts of Respiratory System: The respiratory system consists of the nose, pharynx (throat), larynx (voice box), trachea (windpipe), bronchi and lungs. ASBASJSM College of Pharmacy, Bela, Ropar 1 Chapter 2 Respiratory System 1. Nose: It is only externally visible organ of respiratory system. Nose can be divided into 2 parts: External nose: The external nose consists of a framework of bones and hyaline cartilage covered with muscle and skin and lined by a mucous membrane. The frontal bone, nasal bones and maxillae form the bony framework of the external nose. On the undersurface of the external nose are two openings called the external nares or nostrils. Interior to external nostrils is a portion called nasal vestibule which has coarse hairs that filter out dust particles.