The Nature of Life: Classical and Contemporary Perspectives from Philosophy and Science
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Development, Evolution, and Adaptation Author(S): Kim Sterelny Source: Philosophy of Science, Vol
Philosophy of Science Association Development, Evolution, and Adaptation Author(s): Kim Sterelny Source: Philosophy of Science, Vol. 67, Supplement. Proceedings of the 1998 Biennial Meetings of the Philosophy of Science Association. Part II: Symposia Papers (Sep., 2000), pp. S369-S387 Published by: The University of Chicago Press on behalf of the Philosophy of Science Association Stable URL: http://www.jstor.org/stable/188681 Accessed: 03/03/2010 17:09 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=ucpress. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Philosophy of Science Association and The University of Chicago Press are collaborating with JSTOR to digitize, preserve and extend access to Philosophy of Science. -

Foucault's Darwinian Genealogy
genealogy Article Foucault’s Darwinian Genealogy Marco Solinas Political Philosophy, University of Florence and Deutsches Institut Florenz, Via dei Pecori 1, 50123 Florence, Italy; [email protected] Academic Editor: Philip Kretsedemas Received: 10 March 2017; Accepted: 16 May 2017; Published: 23 May 2017 Abstract: This paper outlines Darwin’s theory of descent with modification in order to show that it is genealogical in a narrow sense, and that from this point of view, it can be understood as one of the basic models and sources—also indirectly via Nietzsche—of Foucault’s conception of genealogy. Therefore, this essay aims to overcome the impression of a strong opposition to Darwin that arises from Foucault’s critique of the “evolutionistic” research of “origin”—understood as Ursprung and not as Entstehung. By highlighting Darwin’s interpretation of the principles of extinction, divergence of character, and of the many complex contingencies and slight modifications in the becoming of species, this essay shows how his genealogical framework demonstrates an affinity, even if only partially, with Foucault’s genealogy. Keywords: Darwin; Foucault; genealogy; natural genealogies; teleology; evolution; extinction; origin; Entstehung; rudimentary organs “Our classifications will come to be, as far as they can be so made, genealogies; and will then truly give what may be called the plan of creation. The rules for classifying will no doubt become simpler when we have a definite object in view. We possess no pedigrees or armorial bearings; and we have to discover and trace the many diverging lines of descent in our natural genealogies, by characters of any kind which have long been inherited. -

The History of Planet Earth
SECOND EDITION Earth’s Evolving The History of Systems Planet Earth Ronald Martin, Ph.D. University of Delaware Newark, Delaware © Jones & Bartlett Learning, LLC, an Ascend Learning Company. NOT FOR SALE OR DISTRIBUTION 9781284457162_FMxx_00i_xxii.indd 1 07/11/16 1:46 pm World Headquarters Jones & Bartlett Learning 5 Wall Street Burlington, MA 01803 978-443-5000 [email protected] www.jblearning.com Jones & Bartlett Learning books and products are available through most bookstores and online booksellers. To contact Jones & Bartlett Learning directly, call 800-832-0034, fax 978-443-8000, or visit our website, www.jblearning.com. Substantial discounts on bulk quantities of Jones & Bartlett Learning publications are available to corporations, professional associations, and other qualified organizations. For details and specific discount information, contact the special sales department at Jones & Bartlett Learning via the above contact information or send an email to [email protected]. Copyright © 2018 by Jones & Bartlett Learning, LLC, an Ascend Learning Company All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without written permission from the copyright owner. The content, statements, views, and opinions herein are the sole expression of the respective authors and not that of Jones & Bartlett Learning, LLC. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement or recommendation by Jones & Bartlett Learning, LLC, and such reference shall not be used for advertis- ing or product endorsement purposes. -
![On Spirituality Century with the Rise of Secular Views on Spirituality [10, 11]](https://docslib.b-cdn.net/cover/5584/on-spirituality-century-with-the-rise-of-secular-views-on-spirituality-10-11-215584.webp)
On Spirituality Century with the Rise of Secular Views on Spirituality [10, 11]
February 2008 The Irish Psychologist ARTICLE historical and conceptual coupling between religiosity and spirituality, a conceptual coupling that has been thrown into disarray over the past On Spirituality century with the rise of secular views on spirituality [10, 11]. I recently asked a group of third year Part 1: Deconstructing, students, "Do you think psychologists should study spirituality?" Some of grappling, and moving in the them said, "No, spirituality cannot be observed or objectively measured – it has no place in psychological field of others science." I wondered if this reply reflected a misunderstanding of Michael J. Hogan the goals of psychological science - a curious negation of certain Mike Hogan is a lecturer in psychology at the National University of Ireland, categories of human experience as a Galway. Correspondence regarding this article may be addressed by email to focus of inquiry - or simply a dislike [email protected]. for entertaining ideas that are in some way associated with orthodox religion. In truth, I cannot claim to have direct The highest activity a human being can and every attempt to construct an access to the minds of my students, attain is learning for understanding, objective account of consciousness and I can only speculate as to why because to understand is to be free struggles with the problem of human they think the way they think. At the Baruch Spinoza, Ethics subjectivity [6, 7]. Consciousness same time, even though the students itself - coupled with our prolific failed to tell me what spirituality is, We are staggeringly lucky to find imaginations - can offer us a sense many students did at least consider ourselves in the spotlight. -

Philosophical Perspectives on Evolutionary Theory
Journal of the Royal Society of Western Australia, 92: 461–464, 2009 Philosophical perspectives on Evolutionary Theory A Tapper Centre for Applied Ethics & Philosophy, Curtin University of Technology, Bentley, WA [email protected] Manuscript received December 2009; accepted February 2010 Abstract Discussion of Darwinian evolutionary theory by philosophers has gone through a number of historical phases, from indifference (in the first hundred years), to criticism (in the 1960s and 70s), to enthusiasm and expansionism (since about 1980). This paper documents these phases and speculates about what, philosophically speaking, underlies them. It concludes with some comments on the present state of the evolutionary debate, where rapid and important changes within evolutionary theory may be passing by unnoticed by philosophers. Keywords: Darwinism, evolutionary theory, philosophy of biology; evolution. Introduction author. Biologists such as Richard Dawkins, Stephen Jay Gould, Steve Jones and Simon Conway Morris are Darwin once said that he had no aptitude for prominent. So also are historians of science, such as Peter philosophy: “My power to follow a long and purely Bowler, Janet Browne, Adrian Desmond and James R. abstract train of thought is very limited; I should, Moore. Equally likely, however, one might be introduced moreover, never have succeeded with metaphysics or to evolutionary theory by a philosopher of biology, for mathematics” (Darwin 1958). This was not false modesty; example Michael Ruse, David Hull or Kim Sterelny. (For -

Nature Based Solutions FEMA
PROMOTING NATURE-BASED HAZARD MITIGATION THROUGH FEMA MITIGATION GRANTS ABBREVIATIONS ADCIRC – Advanced Circulation Model HGM Approach – Hydrogeomorphic Approach BCA – Benefit-Cost Analysis HMA – Hazard Mitigation Assistance BCR – Benefit-Cost Ratio HMGP – Hazard Mitigation Grant Program BRIC – Building Resilient Infrastructure and MSCP – Multiple Species Conservation Program Communities NBS – Nature-Based Solution C&CB – Capability- and Capacity-Building NFIP – National Flood Insurance Program CDBG-DR – Community Development Block Grant- Disaster Recovery NFWF – National Fish and Wildlife Foundation CDBG-MIT – Community Development Block NOAA – National Oceanic and Atmospheric Grant-Mitigation Administration D.C. – District of Columbia NOFO – Notice of Funding Opportunity DEM – Department of Emergency Management NPV – Net Present Value DOI – Department of the Interior SCC – State Coastal Conservancy EDYS – Ecological Dynamics Simulation SDG&E – San Diego Gas & Electric EMA – Emergency Management Agency SFHA – Special Flood Hazard Area EPA SWMM – Environmental Protection Agency SHMO – State Hazard Mitigation Officer Storm Water Management Model SLAMM – Sea Level Affecting Marshes Model FEMA – Federal Emergency Management Agency SRH-2D – Sedimentation and River Hydraulics – FIRM – Flood Insurance Rate Map Two-Dimension FMA – Flood Mitigation Assistance STWAVE – Steady-State Spectral Wave Model FMAG – Fire Management Assistance Grant TNC – The Nature Conservancy HAZUS – Hazards US USACE – U.S. Army Corps of Engineers HEC-HMS – Hydrologic -

Synthesis of Phylogeny and Taxonomy Into a Comprehensive Tree of Life
Synthesis of phylogeny and taxonomy into a comprehensive tree of life Cody E. Hinchliffa,1, Stephen A. Smitha,1,2, James F. Allmanb, J. Gordon Burleighc, Ruchi Chaudharyc, Lyndon M. Coghilld, Keith A. Crandalle, Jiabin Dengc, Bryan T. Drewf, Romina Gazisg, Karl Gudeh, David S. Hibbettg, Laura A. Katzi, H. Dail Laughinghouse IVi, Emily Jane McTavishj, Peter E. Midfordd, Christopher L. Owenc, Richard H. Reed, Jonathan A. Reesk, Douglas E. Soltisc,l, Tiffani Williamsm, and Karen A. Cranstonk,2 aEcology and Evolutionary Biology, University of Michigan, Ann Arbor, MI 48109; bInterrobang Corporation, Wake Forest, NC 27587; cDepartment of Biology, University of Florida, Gainesville, FL 32611; dField Museum of Natural History, Chicago, IL 60605; eComputational Biology Institute, George Washington University, Ashburn, VA 20147; fDepartment of Biology, University of Nebraska-Kearney, Kearney, NE 68849; gDepartment of Biology, Clark University, Worcester, MA 01610; hSchool of Journalism, Michigan State University, East Lansing, MI 48824; iBiological Science, Clark Science Center, Smith College, Northampton, MA 01063; jDepartment of Ecology and Evolutionary Biology, University of Kansas, Lawrence, KS 66045; kNational Evolutionary Synthesis Center, Duke University, Durham, NC 27705; lFlorida Museum of Natural History, University of Florida, Gainesville, FL 32611; and mComputer Science and Engineering, Texas A&M University, College Station, TX 77843 Edited by David M. Hillis, The University of Texas at Austin, Austin, TX, and approved July 28, 2015 (received for review December 3, 2014) Reconstructing the phylogenetic relationships that unite all line- published phylogenies are available only as journal figures, rather ages (the tree of life) is a grand challenge. The paucity of homologous than in electronic formats that can be integrated into databases and character data across disparately related lineages currently renders synthesis methods (7–9). -

The Solar System
5 The Solar System R. Lynne Jones, Steven R. Chesley, Paul A. Abell, Michael E. Brown, Josef Durech,ˇ Yanga R. Fern´andez,Alan W. Harris, Matt J. Holman, Zeljkoˇ Ivezi´c,R. Jedicke, Mikko Kaasalainen, Nathan A. Kaib, Zoran Kneˇzevi´c,Andrea Milani, Alex Parker, Stephen T. Ridgway, David E. Trilling, Bojan Vrˇsnak LSST will provide huge advances in our knowledge of millions of astronomical objects “close to home’”– the small bodies in our Solar System. Previous studies of these small bodies have led to dramatic changes in our understanding of the process of planet formation and evolution, and the relationship between our Solar System and other systems. Beyond providing asteroid targets for space missions or igniting popular interest in observing a new comet or learning about a new distant icy dwarf planet, these small bodies also serve as large populations of “test particles,” recording the dynamical history of the giant planets, revealing the nature of the Solar System impactor population over time, and illustrating the size distributions of planetesimals, which were the building blocks of planets. In this chapter, a brief introduction to the different populations of small bodies in the Solar System (§ 5.1) is followed by a summary of the number of objects of each population that LSST is expected to find (§ 5.2). Some of the Solar System science that LSST will address is presented through the rest of the chapter, starting with the insights into planetary formation and evolution gained through the small body population orbital distributions (§ 5.3). The effects of collisional evolution in the Main Belt and Kuiper Belt are discussed in the next two sections, along with the implications for the determination of the size distribution in the Main Belt (§ 5.4) and possibilities for identifying wide binaries and understanding the environment in the early outer Solar System in § 5.5. -

Ten Misunderstandings About Evolution a Very Brief Guide for the Curious and the Confused by Dr
Ten Misunderstandings About Evolution A Very Brief Guide for the Curious and the Confused By Dr. Mike Webster, Dept. of Neurobiology and Behavior, Cornell Lab of Ornithology, Cornell University ([email protected]); February 2010 The current debate over evolution and “intelligent design” (ID) is being driven by a relatively small group of individuals who object to the theory of evolution for religious reasons. The debate is fueled, though, by misunderstandings on the part of the American public about what evolutionary biology is and what it says. These misunderstandings are exploited by proponents of ID, intentionally or not, and are often echoed in the media. In this booklet I briefly outline and explain 10 of the most common (and serious) misunderstandings. It is impossible to treat each point thoroughly in this limited space; I encourage you to read further on these topics and also by visiting the websites given on the resource sheet. In addition, I am happy to send a somewhat expanded version of this booklet to anybody who is interested – just send me an email to ask for one! What are the misunderstandings? 1. Evolution is progressive improvement of species Evolution, particularly human evolution, is often pictured in textbooks as a string of organisms marching in single file from “simple” organisms (usually a single celled organism or a monkey) on one side of the page and advancing to “complex” organisms on the opposite side of the page (almost invariably a human being). We have all seen this enduring image and likely have some version of it burned into our brains. -
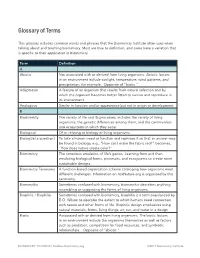
Glossary of Terms
Glossary of Terms This glossary includes common words and phrases that the Biomimicry Institute often uses when talking about and teaching biomimicry. Most are true to definition, and some have a variation that is specific to their application in biomimicry. Term Definition A Abiotic Not associated with or derived from living organisms. Abiotic factors in an environment include sunlight, temperature, wind patterns, and precipitation, for example. Opposite of "biotic." Adaptation A feature of an organism that results from natural selection and by which the organism becomes better fitted to survive and reproduce in its environment. Analogous Similar in function and/or appearance but not in origin or development. B Biodiversity The variety of life and its processes; includes the variety of living organisms, the genetic differences among them, and the communities and ecosystems in which they occur. Biological Of or relating to biology or living organisms. Biologize [a question] To take a human need or function and rephrase it so that an answer may be found in biology, e.g., "How can I make the fabric red?" becomes, "How does nature create color?" Biomimicry The conscious emulation of life’s genius. Learning from and then emulating biological forms, processes, and ecosystems to create more sustainable designs. Biomimicry Taxonomy A function-based organization scheme cataloging how organisms meet different challenges. Information on AskNature.org is organized by this taxonomy. Biomorphic Sometimes confused with biomimicry, biomorphic describes anything resembling or suggesting the forms of living organisms. Biophilic / Biophilia Sometimes confused with biomimicry, biophilia is a term popularized by E.O. Wilson to describe the extent to which humans need connection with nature and other forms of life. -

Darwin's “Tree of Life”
Icons of Evolution? Why Much of What Jonathan Wells Writes about Evolution is Wrong Alan D. Gishlick, National Center for Science Education DARWIN’S “TREE OF LIFE” mon descent. Finally, he demands that text- books treat universal common ancestry as PHYLOGENETIC TREES unproven and refrain from illustrating that n biology, a phylogenetic tree, or phyloge- “theory” with misleading phylogenies. ny, is used to show the genealogic relation- Therefore, according to Wells, textbooks Iships of living things. A phylogeny is not should state that there is no evidence for com- so much evidence for evolution as much as it mon descent and that the most recent research is a codification of data about evolutionary his- refutes the concept entirely. Wells is complete- tory. According to biological evolution, organ- ly wrong on all counts, and his argument is isms share common ancestors; a phylogeny entirely based on misdirection and confusion. shows how organisms are related. The tree of He mixes up these various topics in order to life shows the path evolution took to get to the confuse the reader into thinking that when current diversity of life. It also shows that we combined, they show an endemic failure of can ascertain the genealogy of disparate living evolutionary theory. In effect, Wells plays the organisms. This is evidence for evolution only equivalent of an intellectual shell game, put- in that we can construct such trees at all. If ting so many topics into play that the “ball” of evolution had not happened or common ances- evolution gets lost. try were false, we would not be able to discov- THE CAMBRIAN EXPLOSION er hierarchical branching genealogies for ells claims that the Cambrian organisms (although textbooks do not general- Explosion “presents a serious chal- ly explain this well). -

Sex and Death: an Introduction to Philosophy of Biology Pdf Free
SEX AND DEATH: AN INTRODUCTION TO PHILOSOPHY OF BIOLOGY PDF, EPUB, EBOOK Kim Sterelny,Paul Griffiths | 416 pages | 15 Jun 1999 | The University of Chicago Press | 9780226773049 | English | Chicago, IL, United States Sex and Death: An Introduction to Philosophy of Biology PDF Book Beyond the Modular Emotions Harriet S. The Nature of Species 9. Reduction by Degrees? Griffiths present both the science and the philosophical context necessary for a critical understanding of the most exciting debates shaping biology today. Overall I wouldn't say it is a bad book, and certainly gives tons of information about philosophy of biology, but I think I tried to bite off more than I can chew. Enlightenment, Compassion and Bioscientific Phenomena 4. Replicators and Interactors 3. How can I wanted to dig deeper into evolutionary biology, but I had two problems. There are few things I got from it. Privacy Policy Terms of Use. Other editions. Is the history of life a series of accidents or a drama scripted by selfish genes? However, for technical advancements to make real impacts both on patient health and genuine scientific understanding, quite a number of lingering challenges facing the entire spectrum from protein biology all the way to randomized controlled trials should start to be overcome. Oceanography and Marine Biology. Crandall , Norman C. Reduction in General Philosophy of Science. Images Additional images. Informed answers to questions like these, critical to our understanding of ourselves and the world around us, require both a knowledge o Is the history of life a series of accidents or a drama scripted by selfish genes? The Balance of Nature In this accessible introduction to philosophy of biology, Kim Sterelny and Paul E.