Diversity and Abundance of Cerambycid Beetles&Nbsp
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Updated 02.03.2021 an Annotated List of Afghanistan Longhorn Beetles
Updated 02.03.2021 An annotated list of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) M.A. Lazarev The list is based on the publication by M. A. Lazarev, 2019: Catalogue of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) with two descriptions of new Phytoecia (Parobereina Danilevsky, 2018) from Central Asia. - Humanity space. International almanac, 8 (2): 104-140.It is regularly updated as new taxa are described (marked with red). Family CERAMBYCIDAE Latreille, 1802 subfamily Prioninae Latreille, 1802 tribe Macrotomini J. Thomson, 1861 genus Anomophysis Quentin & Villiers, 1981: 374 type species Prionus spinosus Fabricius, 1787 inscripta C.O. Waterhouse, 1884: 380 (Macrotoma) Heyrovský, 1936: 211 - Wama; Tippmann, 1958: 41 - Kabul, Ost-Afghanistan, 1740; Sarobi, am Kabulflus, 900 m; Mangul, Bashgultal, Nuristan, Ost-Afghanistan, 1250 m; Fuchs, 1961: 259 - Sarobi 1100 m, O.- Afghanistan; Fuchs, 1967: 432 - Afghanistan, 25 km N von Barikot, 1800 m, Nuristan; Nimla, 40 km SW von Dschelalabad; Heyrovský, 1967: 156 - Zentral-Afghanistan, Prov. Kabul: Kabul; Kabul-Zahir; Ost- Afghanistan, Prov. Nengrahar: “Nuristan”, ohne näheren Fundort; Kamu; Löbl & Smetana, 2010: 90 - Afghanistan. plagiata C.O. Waterhouse, 1884: 381 (Macrotoma) vidua Lameere, 1903: 167 (Macrotoma) Quentin & Villiers, 1981: 361, 376, 383 - Afghanistan: Kaboul; Sairobi; Darah-i-Nour, Nangahar; Löbl & Smetana, 2010: 90 - Afghanistan; Kariyanna et al., 2017: 267 - Afghanistan: Kaboul; Rapuzzi et al., 2019: 64 - Afghanistan. tribe Prionini Latreille, 1802 genus Dorysthenes Vigors, 1826: 514 type species Prionus rostratus Fabricius, 1793 subgenus Lophosternus Guérin-Méneville, 1844: 209 type species Lophosternus buquetii Guérin- Méneville, 1844 Cyrtosternus Guérin-Méneville, 1844: 210 type species Lophosternus hopei Guérin-Méneville, 1844 huegelii L. Redtenbacher, 1844: 550 (Cyrtognathus) falco J. -

Jurnal Pendidikan Biologi, Vol 8, No 1, April 2021 INVENTARISASI DAN
87 INVENTARISASI DAN KARAKTERISASI KUMBANG ANTENA PANJANG (CERAMBYCIDAE) DI HUTAN TALANG PEMATANG PUDU PROVINSI RIAU Ennie Chahyadi1), Fahri2), Dilla Asrizalni3) 1),3)Jurusan Biologi, Universitas Riau, 2)Jurusan Biologi, Universitas Tadulako. email1): [email protected] email2): [email protected] email3): [email protected] ABSTRAK: Kumbang longhorn yang dikenal dengan nama kumbang cerambycid merupakan kumbang yang memiliki peran sangat penting dalam proses pembusukan yang berguna bagi keseimbangan ekosistem hutan. Keanekaragaman kumbang ini masih belum banyak diketahui dan dipelajari di Indonesia, terutama di Provinsi Riau. Sedangkan provinsi ini memiliki heterogenitas vegetasi yang tinggi yang merupakan habitat dan sumber makanan bagi kumbang antena panjang. Penelitian ini bertujuan untuk menginventarisasi dan mengkarakterisasi kumbang longhorn di hutan Talang Pematang Pudu Provinsi Riau. Metode yang digunakan adalah menggunakan perangkap Arthrocarpus. Kumbang dikumpulkan pada pukul 15.00 wib. Kemudian dilakukan pengawetan dan identifikasi karakter morfologi berdasarkan ciri kepala, dada dan perut. Hasil penelitian didapatkan sebanyak 12 spesies kumbang longhorn, dengan genera yang terdiri dari Sybra, Pterolophia, dan Epepeotes dari Subfamili Lamiinae. Jumlah individu yang diperoleh sebanyak 110 kumbang. Ciri-ciri kumbang longhorn yang dapat membedakan jenis kumbang terdapat pada ciri-ciri antena, elytra, skutelum dan pronotum. Kata Kunci : Kumbang longhorn, Cerambycid, Riau ABSTRACT:The longhorn beetle known as the Cerambycid beetle, is a beetle that has a very important role in the decomposition process which is useful for the balance of the forest ecosystem. The diversity of these beetles is still not widely known and studied in Indonesia, expectially in Riau Province. Meanwhile, the province has high vegetation heterogeneity which is a habitat and source of food for long antennae beetles. -

Envenomations in Humans Caused by The
linica f C l To o x l ic a o n r l o u g o y J Amaral et al., J Clin Toxicol 2018, 8:4 Journal of Clinical Toxicology DOI: 10.4172/2161-0495.1000392 ISSN: 2161-0495 Case Report Open Access Envenomations in Humans Caused by the Venomous Beetle Onychocerus albitarsis: Observation of Two Cases in São Paulo State, Brazil Amaral ALS1*, Castilho AL1, Borges de Sá AL2 and Haddad V Jr3 1Departamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista – UNESP, CEP 18618-000, Botucatu, São Paulo State, Brazil 2Private Clinic, Botucatu, São Paulo State, Brazil 3Departamento de Dermatologia e Radioterapia, Faculdade de Medicina, Universidade Estadual Paulista – UNESP, CP 557, CEP 18618-000, Botucatu, São Paulo State, Brazil *Corresponding author: Antonio L. Sforcin Amaral, Departamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista – UNESP, CEP 18618-000, Botucatu, São Paulo State, Brazil, Email: [email protected] Received date: July 23, 2018; Accepted date: August 21, 2018; Published date: August 24, 2018 Copyright: ©2018 Amaral ALS, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Abstract Beetles (Coleoptera) are the most diverse group of animals in the world and occur in many environments. In Atlantic and Amazon rainforests, the scorpion-beetle Onychocerus albitarsis (Cerambycidae), can be found. It has venom glandules and inoculators organs in the antenna extremities. Two injuries in humans are reported, showing different patterns of skin reaction after the stings. -

Exploring Flat Faced Longhorn Beetles (Cerambycidae: Lamiinae) from the Reserve Forests of Dooars, West Bengal, India
Hindawi Publishing Corporation ISRN Entomology Volume 2013, Article ID 737193, 8 pages http://dx.doi.org/10.1155/2013/737193 Research Article Exploring Flat Faced Longhorn Beetles (Cerambycidae: Lamiinae) from the Reserve Forests of Dooars, West Bengal, India Sumana Saha,1 Hüseyin Özdikmen,2 Manish Kanti Biswas,3 and Dinendra Raychaudhuri4 1 Department of Zoology, Darjeeling Government College, Government of West Bengal, Darjeeling, West Bengal 734101, India 2 Gazi Universitesi,¨ Fen-Edebiyat Fakultesi,¨ Biyoloji Bol¨ um¨ u,¨ 06500 Ankara, Turkey 3 Department of Zoology, Sreegopal Banerjee College, Mogra, Hooghly, West Bengal 712148, India 4 Entomology Laboratory, Department of Zoology, University of Calcutta, 35 Ballygunge Circular Road, Kolkata, West Bengal 700019, India Correspondence should be addressed to Dinendra Raychaudhuri; [email protected] Received 25 June 2013; Accepted 7 August 2013 Academic Editors: Y. Fan and P. Simoes˜ Copyright © 2013 Sumana Saha et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The present study deals with 29 lamiid species under 21 genera of Dooars, West Bengal, India. These include 4 newly recorded species, namely, Macrochenus isabellinus Aurivillius, Aesopida malasiaca Thomson, Pterolophia (Hylobrotus) lateralis Gahan and Nupserha quadrioculata (Thunberg) from India while 16 others (marked by ∙)fromthestate. 1. Introduction We (saving the second author) for nearly two decades are involved in the exploration of the long horn beetles of Subfamily Lamiinae (Cerambycidae) include members of flat the area. Present communication is one such outcome on the faced longhorn beetles that are both xylophagous and phy- lamiids dealing with 29 species under 21 genera. -

25Th U.S. Department of Agriculture Interagency Research Forum On
US Department of Agriculture Forest FHTET- 2014-01 Service December 2014 On the cover Vincent D’Amico for providing the cover artwork, “…and uphill both ways” CAUTION: PESTICIDES Pesticide Precautionary Statement This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife--if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Product Disclaimer Reference herein to any specific commercial products, processes, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement, recom- mendation, or favoring by the United States government. The views and opinions of wuthors expressed herein do not necessarily reflect those of the United States government, and shall not be used for advertising or product endorsement purposes. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, sexual orientation, or marital or family status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at 202-720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 326-W, Whitten Building, 1400 Independence Avenue, SW, Washington, D.C. -

Catalogue of Afghanistan Longhorn Beetles (Coleoptera, Cerambycidae) with Two Descriptions of New Phytoecia (Parobereina Danilevsky, 2018) from Central Asia
Humanity space International almanac VOL. 8, No 2, 2019: 104-140 http://zoobank.org/urn:lsid:zoobank.org:pub:30F6FA0A-2D7A-4ED2-9EAE-AB7707FFBE61 Catalogue of Afghanistan Longhorn beetles (Coleoptera, Cerambycidae) with two descriptions of new Phytoecia (Parobereina Danilevsky, 2018) from Central Asia M.A. Lazarev State Budget Professional Educational Institution of the Moscow Region “Chekhov technical college” Novaya str., 4, Novyi Byt village, Chekhov District, Moscow Region 142322 Russia e-mail: [email protected]; [email protected] Key words: Coleoptera, Cerambycidae, taxonomy, distribution, new species, Afghanistan, Pakistan. Abstract: The Catalogue includes all 78 Cerambycidae species of Afghanistan fauna known up to 2019 with the references to the original descriptions; 22 species were not mentioned for Afghanistan in Palaearctic Cerambycidae Catalogue by Löbl & Smetana (2010). Bibliography of each species usually includes the geographical information from corresponding publications. Many new taxonomy positions published after 2010 are used here without special remarks. Agapanthia (Epoptes) dahli ustinovi Danilevsky, 2013 stat. nov. is downgraded from the species level. Two species are described as new Phytoecia (Parobereina) pashtunica sp. n. from Afghanistan and Phytoecia (Parobereina) heinzi sp.n. from Pakistan. The present work is an attempt to summarize all data published up to now on Cerambycidae of Afghanistan fauna. Family CERAMBYCIDAE Latreille, 1802 subfamily Prioninae Latreille, 1802 tribe Macrotomini J. Thomson, 1861 genus Anomophysis Quentin & Villiers, 1981: 374 type species Prionus spinosus Fabricius, 1787 inscripta C.O. Waterhouse, 1884: 380 (Macrotoma) Heyrovský, 1936: 211 - Wama; Tippmann, 1958: 41 - Kabul, Ost- Afghanistan, 1740; Sarobi, am Kabulflus, 900 m; Mangul, Bashgultal, Nuristan, Ost-Afghanistan, 1250 m; Fuchs, 1961: 259 - Sarobi 1100 m, O.-Afghanistan; Fuchs, 1967: 432 - Afghanistan, 25 km N von Barikot, 1800 m, Nuristan; Nimla, 40 km SW von Dschelalabad; Heyrovský, 1967: 156 - Zentral-Afghanistan, Prov. -
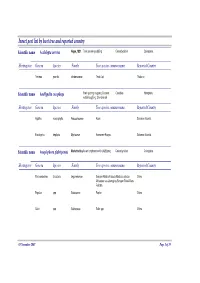
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

Coleoptera: Cerambycidae: Lamiinae: Mesosini
Zoological Studies 51(6): 819-831 (2012) Revision of the Reinstated Genus Pachyosa Fairmaire, 1897 (Coleoptera: Cerambycidae: Lamiinae: Mesosini) Junsuke Yamasako1,* and Nobuo Ohbayashi2 1Entomological Laboratory, Faculty of Agriculture, Ehime University, Tarumi, Matsuyama 790-8566, Japan 2Kamimiyada 1334-444, Minamishitaura-machi, Miura City, 238-0101 Japan (Accepted March 12, 2012 ) Junsuke Yamasako and Nobuo Ohbayashi (2012) Revision of the reinstated genus Pachyosa Fairmaire, 1897 (Coleoptera: Cerambycidae: Lamiinae: Mesosini). Zoological Studies 51(6): 819-831. The genus Pachyosa Fairmaire, 1897, which had been treated as a synonym of Mesosa (Saimia), is reinstated. Five species and 1 subspecies distributed in southwestern Japan and Taiwan are included in the genus as follows: P. cervinopicta Fairmaire, 1897, comb. rev.; P. hirtiventris (Gressitt, 1937), comb. nov.; P. kojimai (Hayashi, 1974), comb. nov.; P. atronotata atronotata (Kusama and Irie, 1976), comb. nov.; P. atronotata yamawakii (Hayashi, 1976), comb. nov.; and P. itoi (N. Ohbayashi, 1985), comb. nov. http://zoolstud.sinica.edu.tw/Journals/51.6/819.pdf Key words: Mesosa (Saimia), Endophallus, Taxonomy, Japan, Taiwan. The monotypic genus Pachyosa was erected Pachyosa, distributed in southwestern Japan by Fairmaire (1897) for P. cervinopicta Fairmaire, and Taiwan, including M. (S.) cervinopicta, are a 1897. Later, Breuning (1938-1940) synonymized homogeneous group, and were considered to be Pachyosa with the subgenus Saimia Pascoe, 1866 an independent genus because of their unique of the genus Mesosa Latreille, 1829. endophallic structures. Therefore, the genus Saimia Pascoe, 1866 was a substitute Pachyosa should be reinstated for this group of name for the genus Samia Pascoe, 1865 (nec Mesosa (Saimia). Herein, we would now like to Hübner, 1819), of which the type species is Samia revise the genus Pachyosa based mainly on the albidorsalis Pascoe, 1865. -

Chapter 10 Insects As
CHAPTER TEN INSECTS 10.1 The insect fauna of south central Seram The insect fauna of Seram is extraordinarily diverse and abundant. Forbes, 1885: 291, writing in his A Naturalist's Wanderings, reports that on Ambon alone insects - particularly beetles - are numerous and of great variety. By the end of the nineteenth century, Ribbe [Ribbe, 1892: 46] had recorded some 19,000 different species, including 10,000 butterflies, and reckoned to collect 300 insects daily. Today the number of certain species for all orders of insects must be many more than this. Clearly, only a very few specimens compared with the total number of known species were col- lected in the field, but even so they constitute the largest single group of specimens. Most importantly they include all common species encountered by the Nuaulu. A checklist of insect species for which specimens were col- lected in south Seram is presented in table 17. 10.2 Nuaulu categories applied to insects Nuaulu terms for insects represent the largest single group in their animal inventory and provide the ethnographer with the greatest problems in presentation and analysis. 10.2.1 makasisi popole The Nuaulu name for this predaceous insect, in AM `capung', relates to its habit of feeding on insects (such as mosquitos) immediately above fresh water, and to the touching of the surface of the water or tips of plants with its tail. Maka comes from makae (`hard'), referring to the head; and sisi, meaning 'to tap, touch or scrape'. Thus eresisi waene is 'to scrape (the) water', where ere is a pronominal vowel prefix indicating a non-human actor. -

Zootaxa, Catalogue of Family-Group Names in Cerambycidae
Zootaxa 2321: 1–80 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Monograph ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) ZOOTAXA 2321 Catalogue of family-group names in Cerambycidae (Coleoptera) YVES BOUSQUET1, DANIEL J. HEFFERN2, PATRICE BOUCHARD1 & EUGENIO H. NEARNS3 1Agriculture and Agri-Food Canada, Central Experimental Farm, Ottawa, Ontario K1A 0C6. E-mail: [email protected]; [email protected] 2 10531 Goldfield Lane, Houston, TX 77064, USA. E-mail: [email protected] 3 Department of Biology, Museum of Southwestern Biology, University of New Mexico, Albuquerque, NM 87131-0001, USA. E-mail: [email protected] Corresponding author: [email protected] Magnolia Press Auckland, New Zealand Accepted by Q. Wang: 2 Dec. 2009; published: 22 Dec. 2009 Yves Bousquet, Daniel J. Heffern, Patrice Bouchard & Eugenio H. Nearns CATALOGUE OF FAMILY-GROUP NAMES IN CERAMBYCIDAE (COLEOPTERA) (Zootaxa 2321) 80 pp.; 30 cm. 22 Dec. 2009 ISBN 978-1-86977-449-3 (paperback) ISBN 978-1-86977-450-9 (Online edition) FIRST PUBLISHED IN 2009 BY Magnolia Press P.O. Box 41-383 Auckland 1346 New Zealand e-mail: [email protected] http://www.mapress.com/zootaxa/ © 2009 Magnolia Press All rights reserved. No part of this publication may be reproduced, stored, transmitted or disseminated, in any form, or by any means, without prior written permission from the publisher, to whom all requests to reproduce copyright material should be directed in writing. This authorization does not extend to any other kind of copying, by any means, in any form, and for any purpose other than private research use. -

(Coleoptera) of Australia
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS McKeown, K. C., 1947. Catalogue of the Cerambycidae (Coleoptera) of Australia. Australian Museum Memoir 10: 1–190. [2 May 1947]. doi:10.3853/j.0067-1967.10.1947.477 ISSN 0067-1967 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia THE AUSTRALIAN MUSEUM, SYDNEY MEMOIR X. CATALOGUE OF THE CERAMBYCIDAE (COLEOPTERA) OF AUSTRALIA BY KEITH C. McKEOWN, F.R.Z.S., Assistant Entomologist. The Australian Museum. PUBLISHED BY ORDER OF THE TRUSTEES A. B. Walkom, D.%., Director. Sydney, May 2, I947 PREFACE. The accompanying Catalogue of the Cerambycidae is the first, dealing solely with Australian genera and species, to be published since that of Pascoe in 1867. Masters' Catalogue of the Described Coleoptera of Australia, 1885-1887, included the Cerambycidae, and was based on the work of Gemminger and Harold. A new catalogue has been badly needed owing to the large number of new species described in recent years, and the changes in the already complicated synonymy. The Junk catalogue, covering the Coleoptera of the world, is defective in many respects, as well as being too unwieldy, and too costly for the average Australian worker. Many of the references in the Junk catalogue are inaccurate, synonymy misleading, and the genera under which the species were originally described omitted, and type localities are not quoted. In this catalogue every care has been taken to ensure accuracy, and the fact that it has been used, in slip form, over a number. -

Coleoptera: Cerambycidae) of Assam, India
Rec. zool. Surv. India: Vol. 117(1)/ 78-90, 2017 ISSN (Online) : (Applied for) DOI: 10.26515/rzsi/v117/i1/2017/117286 ISSN (Print) : 0375-1511 An updated list of cerambycid beetles (Coleoptera: Cerambycidae) of Assam, India Bulganin Mitra1*, Udipta Chakraborti1, Kaushik Mallick1, Subhrajit Bhaumik2 and Priyanka Das1 1Zoological Survey of India, Prani Vigyan Bhavan, M-Block, New Alipore, Kolkata – 700 053, West Bengal, India; [email protected] 2Post Graduate, Department of Zoology, Vidyasagar College, Kolkata – 700006, West Bengal, India Abstract consolidated updated list of cerambycid fauna of Assam and reports 95 species, 64 genera, 32 tribes and 3 subfamilies. AmongAssam isthe a threestate subfamiliesin North-East from India Assam, which subfamily is considered Lamiinae as shares a biological 49 species, hotspot. followed Present by the communication subfamily Cerambycinae is the first with 38 species and Prioninae with only 8 species. Keywords: Longhorn beetle, Assam, North-East India Introduction world, therefore this beetle family is considered as one of important coleopteran family (Agarwala & Bhattacharjee, The study on long horned beetles from the northeast 2012). This communication is the first updated Indian state Assam is very poor with many species consolidated list of cerambycid beetles from the state of awaiting discovery, study and description. Among the Assam (after complete separation from other states of NE seven sister states, cerambycid fauna of Arunachal India in 1987) which includes 95 species under 64 genera Pradesh, Tripura, Meghalaya, Manipur, Mizoram, of 32 tribes belonging to 3 subfamilies along with their Nagaland are mostly worked out by the Zoological Survey distribution. of India and some other universities and institutions.