Tese Doutorado Lorena Christine Ferreira Da Silva.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chapitre Quatre La Spécificité D'hôtes Des Virophages Sputnik
AIX-MARSEILLE UNIVERSITE FACULTE DE MEDECINE DE MARSEILLE ECOLE DOCTORALE DES SCIENCES DE LA VIE ET DE LA SANTE THESE DE DOCTORAT Présentée par Morgan GAÏA Né le 24 Octobre 1987 à Aubagne, France Pour obtenir le grade de DOCTEUR de l’UNIVERSITE AIX -MARSEILLE SPECIALITE : Pathologie Humaine, Maladies Infectieuses Les virophages de Mimiviridae The Mimiviridae virophages Présentée et publiquement soutenue devant la FACULTE DE MEDECINE de MARSEILLE le 10 décembre 2013 Membres du jury de la thèse : Pr. Bernard La Scola Directeur de thèse Pr. Jean -Marc Rolain Président du jury Pr. Bruno Pozzetto Rapporteur Dr. Hervé Lecoq Rapporteur Faculté de Médecine, 13385 Marseille Cedex 05, France URMITE, UM63, CNRS 7278, IRD 198, Inserm 1095 Directeur : Pr. Didier RAOULT Avant-propos Le format de présentation de cette thèse correspond à une recommandation de la spécialité Maladies Infectieuses et Microbiologie, à l’intérieur du Master des Sciences de la Vie et de la Santé qui dépend de l’Ecole Doctorale des Sciences de la Vie de Marseille. Le candidat est amené à respecter des règles qui lui sont imposées et qui comportent un format de thèse utilisé dans le Nord de l’Europe permettant un meilleur rangement que les thèses traditionnelles. Par ailleurs, la partie introduction et bibliographie est remplacée par une revue envoyée dans un journal afin de permettre une évaluation extérieure de la qualité de la revue et de permettre à l’étudiant de commencer le plus tôt possible une bibliographie exhaustive sur le domaine de cette thèse. Par ailleurs, la thèse est présentée sur article publié, accepté ou soumis associé d’un bref commentaire donnant le sens général du travail. -

Genome of Phaeocystis Globosa Virus Pgv-16T Highlights the Common Ancestry of the Largest Known DNA Viruses Infecting Eukaryotes
Genome of Phaeocystis globosa virus PgV-16T highlights the common ancestry of the largest known DNA viruses infecting eukaryotes Sebastien Santinia, Sandra Jeudya, Julia Bartolia, Olivier Poirota, Magali Lescota, Chantal Abergela, Valérie Barbeb, K. Eric Wommackc, Anna A. M. Noordeloosd, Corina P. D. Brussaardd,e,1, and Jean-Michel Claveriea,f,1 aStructural and Genomic Information Laboratory, Unité Mixte de Recherche 7256, Centre National de la Recherche Scientifique, Aix-Marseille Université, 13288 Marseille Cedex 9, France; bCommissariat à l’Energie Atomique–Institut de Génomique, 91057 Evry Cedex, France; cDepartment of Plant and Soil Sciences, University of Delaware, Newark, DE 19711; dDepartment of Biological Oceanography, Royal Netherlands Institute for Sea Research, NL-1790 AB Den Burg (Texel), The Netherlands; eAquatic Microbiology, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Amsterdam, The Netherlands; and fService de Santé Publique et d’Information Médicale, Hôpital de la Timone, Assistance Publique–Hôpitaux de Marseille, FR-13385 Marseille, France Edited by James L. Van Etten, University of Nebraska, Lincoln, NE, and approved May 1, 2013 (received for review February 22, 2013) Large dsDNA viruses are involved in the population control of many viruses: 730 kb and 1.28 Mb for CroV and Megavirus chilensis, globally distributed species of eukaryotic phytoplankton and have respectively. Other studies, targeting virus-specific genes [e.g., a prominent role in bloom termination. The genus Phaeocystis (Hap- DNA polymerase B (8) or capsid proteins (9)] have suggested tophyta, Prymnesiophyceae) includes several high-biomass-forming a close phylogenetic relationship between Mimivirus and several phytoplankton species, such as Phaeocystis globosa, the blooms of giant dsDNA viruses infecting various unicellular algae such as which occur mostly in the coastal zone of the North Atlantic and the Pyramimonas orientalis (Chlorophyta, Prasinophyceae), Phaeocys- North Sea. -

Plasmodium Falciparum Is Not As Lonely As Previously Considered
AUTOPHAGIC PUNCTUM ARTICLE ADDENDUM Virulence 2:1, 71-76; January/February 2011; © 2011 Landes Bioscience Plasmodium falciparum is not as lonely as previously considered Franck Prugnolle,1,* Francisco Ayala,2 Benjamin Ollomo,3 Céline Arnathau,1 Patrick Durand1 and François Renaud1,* 1Laboratoire MIVEGEC; UM1-CNRS 5290-IRD 224, IRD Montpellier, France; 2Department of Ecology and Evolutionary Biology; University of California; Irvine, CA USA; 3Centre International de Recherches Médicales de Franceville; Franceville, Gabon ntil very recently, only one species The identification of Plasmodium spe- U(P. reichenowi) was known to be a cies circulating in great apes in Africa phylogenetic sister lineage of P. falciparum, was primarily done during the first half the main malignant agent of human of the twentieth century, on the basis of malaria. In 2009 and 2010, new studies morphological features.1 This approach have revealed the existence of several new has several limitations.4 First, phenotypic phylogenetic species related to this deadly plasticity can lead to incorrect identifica- parasite and infecting chimpanzees and tions. Second, morphological keys are gorillas in Africa. These discoveries invite often effective only for a particular life us to explore a whole set of new questions, stage which cannot always be observed which we briefly do in this article. or is difficult to be. Finally, and perhaps most important, this approach overlooks The Plasmodium species infecting morphologically cryptic taxa. These limi- humans and non-human primates cluster tations, together with the difficulty to into two distinct phylogenetic lineages collect and manipulate great apes, were (Fig. 1). One of these lineages (in yellow certainly, at least in part, responsible for in Fig. -

Legionella Genus Genome Provide Multiple, Independent Combinations for Replication in Human Cells
Supplemental Material More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells Laura Gomez-Valero1,2, Christophe Rusniok1,2, Danielle Carson3, Sonia Mondino1,2, Ana Elena Pérez-Cobas1,2, Monica Rolando1,2, Shivani Pasricha4, Sandra Reuter5+, Jasmin Demirtas1,2, Johannes Crumbach1,2, Stephane Descorps-Declere6, Elizabeth L. Hartland4,7,8, Sophie Jarraud9, Gordon Dougan5, Gunnar N. Schroeder3,10, Gad Frankel3, and Carmen Buchrieser1,2,* Table S1: Legionella strains analyzed in the present study Table S2: Type IV secretion systems predicted in the genomes analyzed Table S3: Eukaryotic like domains identified in the Legionella proteins analyzed Table S4: Small GTPases domains detected in the genus Legionella as defined in the CDD ncbi domain database Table S5: Eukaryotic like proteins detected in the Legionella genomes analyzed in this study Table S6: Aminoacid identity of the Dot/Icm components in Legionella species with respect to orthologous proteins in L. pneumophila Paris Table S7: Distribution of seventeen highly conserved Dot/Icm secreted substrates Table S8: Comparison of the effector reperotoire among strains of the same Legionella species Table S9. Number of Dot/Icm secreted proteins predicted in each strain analyzed Table S10: Replication capacity of the different Legionella species analyzed in this study and collection of literature data on Legionella replication Table S11: Orthologous table for all genes of the 80 analyzed strains based on PanOCT. The orthologoss where defined with the program PanOCT using the parameters previously indicated in material and methods.) Figure S1: Distribution of the genes predicted to encode for the biosynthesis of flagella among all Legionella species. -

Ana Paula Fernandes Viana Furtado Avaliação De Alternativas Para
Universidade Federal de Itajubá Instituto de Ciências Puras e Aplicadas Programa de Pós-Graduação em Gestão e Regulação de Recursos Hídricos PROFÁGUA Ana Paula Fernandes Viana Furtado Avaliação de Alternativas para Reabilitação de Lagos Urbanos: O Caso da Lagoa da Pampulha, em Belo Horizonte – MG Itabira- Minas Gerais 2020 Ana Paula Fernandes Viana Furtado Avaliação de Alternativas para Reabilitação de Lagos Urbanos: O Caso da Lagoa da Pampulha, em Belo Horizonte – MG Dissertação apresentada, como requisito parcial para obtenção do título de Mestre, ao Programa de Pós-Graduação em Gestão e Regulação de Recursos Hídricos, Curso de Mestrado Profissional em Rede Nacional em Gestão e Regulação de Recursos Hídricos (PROFÁGUA), na Universidade Federal de Itajubá. Área de concentração: Regulação e Governança de Recursos Hídricos Aprovada em 23 de setembro de 2020. Banca Examinadora: ___________________________________________________ Prof. Dr. Roberto Cezar de Almeida Monte Mor – Orientador Instituto de Ciências Puras e Aplicadas – UNIFEI ____________________________________________________ Prof. Dr. Eduardo de Aguiar do Couto – Co Orientador Instituto de Ciências Puras e Aplicadas – UNIFEI ____________________________________________________ Prof. Dr. Anderson de Assis Morais – Avaliador Interno Instituto de Ciências Puras e Aplicadas – UNIFEI ____________________________________________________ Profª. Drª. Talita Fernanda das Graças Silva – Avaliadora Extena Departamento de Eng. Hidráulica e Recursos Hídricos – UFMG Itabira – Minas Gerais -

Bacterial Species Library with Validly Published Names (Green Curve); and Sequenced Viral Genomes (Blue)
17 March 2014 ESCMID-ESCAR PGEC MARSEILLE «Ignorance and blindness for a post-modern microbiology ! » © by author ESCMID Online Lecture Library Didier Raoult Marseille - France [email protected] www.mediterranee-infection.com © by author « ESCMIDPostmodern scienceOnline makes Lecture the theory Library of its own evolution as discontinuous, catastrophic, nonrectifiable, paradoxical.» P.97 2 INFECTIONS IN THE XXIST CENTURY Infections in the world : 17 million deaths - 30% - The big 3s : HIV - TB – malaria : no vaccine - Respiratory infections : Etiology? - Digestive infections : Etiology? - Vaccine-prevented infections© by : authorAdhesion - Nosocomial infections : Circuits? - EmergingESCMID infections Online : LectureSource Library - Cancer : Communicable 3 IGNORANCE that decreases but reveals ou ARROGANCE BLINDNES that persists and leads to FALSE DEDUCTIONS © by author and UNVERIFIED PREDICTIONSESCMID Online Lecture Library 4 EMERGING DISEASES IGNORANCE: Major and unique acceleration of knowledge in history Blindness False deductions© by author ESCMID Unverified Online predictions Lecture Library So what ? 5 Viruses: Essential Agents of Life by Günther Witzany © by author ESCMID Online Lecture Library p.64 By F.Rohwer 6 Viruses: Essential Agents of Life by Günther Witzany © by author ESCMID Online Lecture Library p.64 By F.Rohwer 7 PROGRESSES MADE IN MICROBIOLOGY FROM 1979 TO 2012 THANKS TO THE DEVELOPMENT OF NEW TECHNOLOGIES Cultured Sequenced © by author a) the ESCMIDleft ordinate axis refers toOnline the cumulative numbers -
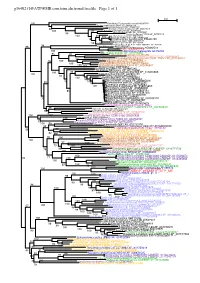
Page 1 G16482.T1@[email protected]
g16482.t1@[email protected] Page 1 of 1 0.2 100 Candidatus Thiomargarita nelsonii KHD07353 Bacillus thuringiensis Bt407 YP_006926419 Legionella pneumophila WP_014842957 88 Endozoicomonas montiporae WP_034879478 Acinetobacter baumannii WP_031968362 100 87 Vibrio nigripulchritudo WP_004402447 98 Psychrobacter sp. P11G3 WP_057760119 100 99 Xenorhabdus bovienii WP_038203911 100 Rouxiella chamberiensis WP_045046329 18 Dendroctonus ponderosae ERL83390 Serratia sp. M24T3 WP_009636677 98 56 100Lonsdalea quercina WP_026743324 Escherichia coli str. K-12 substr. MG1655 NP_415996 86 98 Chlorobi bacterium OLB5 KXK50071 CoptotermesTric_AAA92714 formosanus AGM32510 100 Histomonas meleagridis ACI16482 100Eucalyptus grandis XP_010033838 95 98 Ectocarpus siliculosus CBJ25596 Guillardia theta CCMP2712 XP_005838347 Phaeodactylum tricornutum CCAP 1055/1 XP_002180331 99 Aphanomyces astaci XP_009825348 95 100 Albugo laibachii Nc14 CCA15917 100 Phytophthora parasitica P1569 ETI54450 100 Phytophthora infestans T30-4 XP_002896607 100 Brugia malayi CDP93853 Bombus impatiens XP_003487345 Trichoplax adhaerens XP_002116081 95 Apteryx australis mantelli XP_013810665 100 100 Callorhinchus milii XP_007891153 Homo sapiens AAC50613 93 Callorhinchus milii XP_007900551 100 Clupea harengus XP_012674444 99 Chrysochloris asiatica XP_006865016 82 76 100 84Nannospalax galili XP_008842357 98 Cricetulus griseus XP_007626683 100 65Rousettus aegyptiacus XP_016015809 Condylura cristata XP_004683330 58Chinchilla lanigera XP_013367523 85 61Microcebus murinus XP_012594174 -

And Thermo-Adaptation in Hyperthermophilic Archaea: Identification of Compatible Solutes, Accumulation Profiles, and Biosynthetic Routes in Archaeoglobus Spp
Universidade Nova de Lisboa Osmo- andInstituto thermo de Tecnologia-adaptation Química e Biológica in hyperthermophilic Archaea: Subtitle Subtitle Luís Pedro Gafeira Gonçalves Osmo- and thermo-adaptation in hyperthermophilic Archaea: identification of compatible solutes, accumulation profiles, and biosynthetic routes in Archaeoglobus spp. OH OH OH CDP c c c - CMP O O - PPi O3P P CTP O O O OH OH OH OH OH OH O- C C C O P O O P i Dissertation presented to obtain the Ph.D degree in BiochemistryO O- Instituto de Tecnologia Química e Biológica | Universidade Nova de LisboaP OH O O OH OH OH Oeiras, Luís Pedro Gafeira Gonçalves January, 2008 2008 Universidade Nova de Lisboa Instituto de Tecnologia Química e Biológica Osmo- and thermo-adaptation in hyperthermophilic Archaea: identification of compatible solutes, accumulation profiles, and biosynthetic routes in Archaeoglobus spp. This dissertation was presented to obtain a Ph. D. degree in Biochemistry at the Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa. By Luís Pedro Gafeira Gonçalves Supervised by Prof. Dr. Helena Santos Oeiras, January, 2008 Apoio financeiro da Fundação para a Ciência e Tecnologia (POCI 2010 – Formação Avançada para a Ciência – Medida IV.3) e FSE no âmbito do Quadro Comunitário de apoio, Bolsa de Doutoramento com a referência SFRH / BD / 5076 / 2001. ii ACKNOWNLEDGMENTS The work presented in this thesis, would not have been possible without the help, in terms of time and knowledge, of many people, to whom I am extremely grateful. Firstly and mostly, I need to thank my supervisor, Prof. Helena Santos, for her way of thinking science, her knowledge, her rigorous criticism, and her commitment to science. -

Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere
viruses Review Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere Clara Rolland 1, Julien Andreani 1, Amina Cherif Louazani 1, Sarah Aherfi 1,3, Rania Francis 1 , 1,2 1, 1 1 Rodrigo Rodrigues , Ludmila Santos Silva y, Dehia Sahmi , Said Mougari , 1 1, 1,2 1,§ 1, Nisrine Chelkha , Meriem Bekliz z, Lorena Silva , Felipe Assis ,Fábio Dornas k, Jacques Yaacoub Bou Khalil 3, Isabelle Pagnier 1,3, Christelle Desnues 1,¶, Anthony Levasseur 1,3, Philippe Colson 1,3,Jônatas Abrahão 1,2 and Bernard La Scola 1,3,* 1 MEPHI, APHM, IRD 198, Aix Marseille Univ, Department of Medicine, IHU-Méditerranée Infection, 13005 Marseille, France; [email protected] (C.R.); [email protected] (J.A.); cherifl[email protected] (A.C.L.); aherfi[email protected] (S.A.); [email protected] (R.F.); [email protected] (R.R.); [email protected] (L.S.S.); [email protected] (D.S.); [email protected] (S.M.); [email protected] (N.C.); [email protected] (M.B.); [email protected] (L.S.); [email protected] (F.A.); [email protected] (F.D.); [email protected] (I.P.); [email protected] (C.D.); [email protected] (A.L.); [email protected] (P.C.); [email protected] (J.A.) 2 Laboratório de Vírus, Instituto de Ciêncas Biológicas, Departamento de Microbiologia, Universidade Federal de Minas Gerais, 31270-901 Belo Horizonte, Brazil 3 IHU IHU-Méditerranée Infection, 13005 Marseille, France; [email protected] * Correspondence: [email protected] Present address: Institut de Biologie Moléculaire et Cellulaire, CNRS UPR9022, Université de Strasbourg, y 67000 Strasbourg, France. -

Inferring Natural Selection Signals in Plasmodium Vivax-Encoded Proteins Having a Potential Role in Merozoite Invasion
Identificación de señales de selección natural en genes de Plasmodium vivax que codifican proteínas involucradas en el proceso de invasión para determinar su potencial uso en una vacuna antimalárica. Diego Edison Garzón Ospina Tesis Doctoral presentada como requisito para optar al título de Doctor en Ciencias Biomédicas y Biológicas de la Universidad del Rosario Bogotá, 2018 1 Identificación de señales de selección natural en genes de Plasmodium vivax que codifican proteínas involucradas en el proceso de invasión para determinar su potencial uso en una vacuna antimalárica. Estudiante Diego Edison Garzón Ospina Biólogo, Universidad INCCA de Colombia Magister en Ciencias-Microbiología, Universidad Nacional de Colombia Director Manuel Alfonso Patarroyo Gutiérrez M.D., Dr.Sc. Jefe del Departamento de Biología Molecular e Inmunología Fundación Instituto de Inmunología de Colombia (FIDIC) Profesor Titular, Escuela de Medicina y Ciencias de la Salud Universidad del Rosario DOCTORADO EN CIENCIAS BIOMÉDICAS Y BIOLÓGICAS UNIVERSIDAD DEL ROSARIO Bogotá, 2018 2 AGRADECIMIENTOS Quiero dar mis agradecimientos y dedicar este trabajo a mi madre: Martha Ospina Vargas, y a mis hermanos: Yamile Garzón Ospina y Julián David Escobar Ospina, quienes me han acompañado y apoyado en todo momento. A Sindy Paola Buitrago Puentes por su acompañamiento y apoyo durante este tiempo. También agradecer al Dr. Manuel Alfonso Patarroyo Gutiérrez, por haberme acogido en su equipo de trabajo y por apoyarme a lo largo de todos estos años. Quiero agradecer y reconocer la labor de: Andrea Estefanía Ramos, Darwin Andrés Moreno Pérez, Elizabeth Gutiérrez Vásquez, Heidy Daniela Ortiz Suarez, Lady Johanna Forero Rodríguez, Laura Alejandra Ricaurte Contreras, Leidy Paola Reyes, Luis Alfredo Baquero, Paola Andrea Camargo Ayala, Sindy Paola Buitrago Puentes, Ricardo De León Montero y Yimara Grosso Paz, quienes aportaron su tiempo, dedicación y esfuerzo, permitiendo la culminación de este trabajo. -

Festival Architettura Magazine. Saggi Su Architettura E Città
© FAmagazine - ISSN 2039-0491 DOI: 10.12838/issn.20390491/n25.2014/1 25 - gen-feb/jan-feb 2014 Guilherme Wisnik TRA “DOLCE VITA” E BETWEEN THE “DOLCE SINISTRA VITA” AND THE LEFT Abstract Abstract Morto a quasi 105 anni, Niemeyer ha attraversato mo- Died at almost 105 years, Niemeyer has expe- menti storici cruciali per la storia recente del Brasile rienced important times in the recent history of entrando a pieno titolo a farne parte. Collaboratore Brazil. Collaborator of Lucio Costa and fellow of di Lucio Costa e sodale del presidente Kubitschek, President Kubitschek, the carioca architect has l'architetto carioca ha contribuito a costruire, nel vero helped contributed in building essential parts of senso della parola, parti imprescindibili del paese, the country, making his language and his con- portando il suo linguaggio e la sua concezione pro- ception of the project to be not only an emblem of gettuale ad essere non soltanto emblema nazionale national development for a country with a relatively di sviluppo, per un Paese con una storia relativamen- short history, but also a symbol of the country itself te breve, ma anche simbolo del paese stesso nel in the world. mondo. Born on the 15 December 1907, Oscar Niemeyer Nato il 15 dicembre del 1907, Oscar Niemeyer è mor- died just ten days before celebrating his one hun- to dieci giorni prima di compiere centocinque anni. dred and fifth birthday. Notable longevity, above all if Notevole longevità la sua, soprattutto se osservata observed in parallel with the “youth” of the Brazilian parallelamente alla “giovinezza” della nazione Bra- nation, discovered by the Portuguese in 1500 and sile, scoperto dai portoghesi nel 1500 e proclamato only proclaimed independent in 1822. -

Chapter Four Emergence Re-Evaluated
CHAPTER FOUR EMERGENCE RE-EVALUATED 4.1 INTRODUCTION Whether this will lead to a rejuvenation of a flagging tradition or whether the historical programme of actively developing an “Afrikaner culture” was in any way cardinal to the emergence of a Pretoria Regionalism so as to deny it a continuity as a Third Vernacular has yet to be seen (Fisher 1998:140) He [Fisher] later introduces a surprisingly apt phrase derived from Frampton which has general application: “Modern tempered by critical regionalism”. He introduces too, the concepts of identity and “sentiment”, an 18C term, like the word sublime, which alerts us to the fact that we are in the presence of a myth – in the 19C, the myth of utopia; in the 20C the myth or myths of nationalism and the misreading, if not deliberate falsification, of history (Chipkin 2008:377). This chapter: Re-evaluates the accessible historical continuum and stylistic labels assigned to the ‘emergence’ of our generation. Re-thinks the local stylistic context and situation pertinent to our agents. 4.2 SELECTED LITERATURE This chapter selects literature to investigate the third sub question: What were the local stylistic constructs that formulated an emergence relative to our generation? The introductory quotes imply a literary opposition surrounding regional labelling which sources tie to our generation. In the book he co-edits, Architecture of the Transvaal, Roger Fisher’s essay ‘The Third Vernacular: Pretoria Regionalism – Aspects of an Emergence’ is the first essential source for the critical discourse analysis required for this chapter. If Fisher (1998: 124) postulates, “Afrikaner origins seem to have been pivotal in the emergence of a Pretoria Regionalism”, then Herman Giliomee’s The Afrikaners: Biography of a People (2003) is the most recognised and viable source to assist in a critical re- evaluation of that hypothesis.