Revisiting the Elusive Hepatitis C Vaccine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Contrasting Influenza and Respiratory Syncytial Virus
REVIEW published: 02 March 2018 doi: 10.3389/fimmu.2018.00323 Induction and Subversion of Human Protective Immunity: Contrasting influenza and Respiratory Syncytial Virus Stephanie Ascough, Suzanna Paterson and Christopher Chiu* Section of Infectious Diseases and Immunity, Imperial College London, London, United Kingdom Respiratory syncytial virus (RSV) and influenza are among the most important causes of severe respiratory disease worldwide. Despite the clinical need, barriers to devel- oping reliably effective vaccines against these viruses have remained firmly in place for decades. Overcoming these hurdles requires better understanding of human immunity and the strategies by which these pathogens evade it. Although superficially similar, the virology and host response to RSV and influenza are strikingly distinct. Influenza induces robust strain-specific immunity following natural infection, although protection by current vaccines is short-lived. In contrast, even strain-specific protection is incomplete after RSV Edited by: and there are currently no licensed RSV vaccines. Although animal models have been Steven Varga, critical for developing a fundamental understanding of antiviral immunity, extrapolating University of Iowa, United States to human disease has been problematic. It is only with recent translational advances Reviewed by: (such as controlled human infection models and high-dimensional technologies) that the Tara Marlene Strutt, mechanisms responsible for differences in protection against RSV compared to influenza University of Central Florida, have begun to be elucidated in the human context. Influenza infection elicits high-affinity United States Jie Sun, IgA in the respiratory tract and virus-specific IgG, which correlates with protection. Long- Mayo Clinic Minnesota, lived influenza-specific T cells have also been shown to ameliorate disease. -

Viral Hepatitis Testing Effective Date: January 1, 2012
Viral Hepatitis Testing Effective Date: January 1, 2012 Scope This guideline provides guidance for the use of laboratory tests to diagnose acute and chronic viral hepatitis in adults (> 19 years) in the primary care setting. General Considerations for Ordering Laboratory Tests Prior to ordering tests for hepatitis, consider the patient’s history, age, risk factors (see below), hepatitis vaccination status, and any available previous hepatitis test results. Risk Factors for Viral Hepatitis include: • Substance use (includes sharing drug snorting, smoking or injection equipment) • High-risk sexual activity or sexual partner with viral hepatitis • Travel to or from high-risk hepatitis endemic areas or exposure during a local outbreak • Immigration from hepatitis B and/or C endemic countries • Household contact with an infected person especially if personal items (e.g., razors, toothbrushes, nail clippers) are shared • Recipient of unscreened blood products* • Needle-stick injury or other occupational exposure (e.g., healthcare workers) • Children born to mothers with chronic hepatitis B or C infection • Attendance at daycare • Contaminated food or water (hepatitis A only) • Tattoos and body piercing • History of incarceration • HIV or other sexually transmitted infection • Hemodialysis *screening of donated blood products for hepatitis C (anti-HCV) began in 1990 in Canada.1 Types of Viral Hepatitis Hepatitis A: causes acute but not chronic hepatitis Hepatitis B: causes acute and chronic hepatitis Hepatitis C: causes chronic hepatitis but rarely manifests as acute hepatitis Hepatitis D: rare and only occurs in patients infected with hepatitis B Hepatitis E: clinically similar to hepatitis A, mostly restricted to endemic areas and occasionally causes chronic infection in immunosuppressed people Others: e.g. -

Prevention & Control of Viral Hepatitis Infection
Prevention & Control of Viral Hepatitis Infection: A Strategy for Global Action © World Health Organization 2011. All rights reserved. The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement. The mention of specific companies or of certain manufacturers’ products does not imply that they are endorsed or recommended by the World Health Organization in preference to others of a similar nature that are not mentioned. Errors and omissions excepted, the names of proprietary products are distinguished by initial capital letters. All reasonable precautions have been taken by WHO to verify the information contained in this publication. However, the published material is being distributed without warranty of any kind, either express or implied. The responsibility for the interpretation and use of the material lies with the reader. In no event shall the World Health Organization be liable for damages arising from its use. Table of contents Disease burden 02 What is viral hepatitis? 05 Prevention & control: a tailored approach 06 Global Achievements 08 Remaining challenges 10 World Health Assembly: a mandate for comprehensive prevention & control 13 WHO goals and strategy -

The Role of Hepatitis C Virus in Hepatocellular Carcinoma U
Viruses in cancer cell plasticity: the role of hepatitis C virus in hepatocellular carcinoma U. Hibner, D. Gregoire To cite this version: U. Hibner, D. Gregoire. Viruses in cancer cell plasticity: the role of hepatitis C virus in hepato- cellular carcinoma. Contemporary Oncology, Termedia Publishing House, 2015, 19 (1A), pp.A62–7. 10.5114/wo.2014.47132. hal-02187396 HAL Id: hal-02187396 https://hal.archives-ouvertes.fr/hal-02187396 Submitted on 2 Jun 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution - NonCommercial - ShareAlike| 4.0 International License Review Viruses are considered as causative agents of a significant proportion of human cancers. While the very Viruses in cancer cell plasticity: stringent criteria used for their clas- sification probably lead to an under- estimation, only six human viruses the role of hepatitis C virus are currently classified as oncogenic. In this review we give a brief histor- in hepatocellular carcinoma ical account of the discovery of on- cogenic viruses and then analyse the mechanisms underlying the infectious causes of cancer. We discuss viral strategies that evolved to ensure vi- Urszula Hibner1,2,3, Damien Grégoire1,2,3 rus propagation and spread can alter cellular homeostasis in a way that increases the probability of oncogen- 1Institut de Génétique Moléculaire de Montpellier, CNRS, UMR 5535, Montpellier, France ic transformation and acquisition of 2Université Montpellier 2, Montpellier, France stem cell phenotype. -

The Use of Non-Human Primates in Research in Primates Non-Human of Use The
The use of non-human primates in research The use of non-human primates in research A working group report chaired by Sir David Weatherall FRS FMedSci Report sponsored by: Academy of Medical Sciences Medical Research Council The Royal Society Wellcome Trust 10 Carlton House Terrace 20 Park Crescent 6-9 Carlton House Terrace 215 Euston Road London, SW1Y 5AH London, W1B 1AL London, SW1Y 5AG London, NW1 2BE December 2006 December Tel: +44(0)20 7969 5288 Tel: +44(0)20 7636 5422 Tel: +44(0)20 7451 2590 Tel: +44(0)20 7611 8888 Fax: +44(0)20 7969 5298 Fax: +44(0)20 7436 6179 Fax: +44(0)20 7451 2692 Fax: +44(0)20 7611 8545 Email: E-mail: E-mail: E-mail: [email protected] [email protected] [email protected] [email protected] Web: www.acmedsci.ac.uk Web: www.mrc.ac.uk Web: www.royalsoc.ac.uk Web: www.wellcome.ac.uk December 2006 The use of non-human primates in research A working group report chaired by Sir David Weatheall FRS FMedSci December 2006 Sponsors’ statement The use of non-human primates continues to be one the most contentious areas of biological and medical research. The publication of this independent report into the scientific basis for the past, current and future role of non-human primates in research is both a necessary and timely contribution to the debate. We emphasise that members of the working group have worked independently of the four sponsoring organisations. Our organisations did not provide input into the report’s content, conclusions or recommendations. -

UC Irvine UC Irvine Electronic Theses and Dissertations
UC Irvine UC Irvine Electronic Theses and Dissertations Title Computation Models of Virus Dynamics Permalink https://escholarship.org/uc/item/3zb6480f Author Roy, Sarah M. Publication Date 2015 Peer reviewed|Thesis/dissertation eScholarship.org Powered by the California Digital Library University of California UNIVERSITY OF CALIFORNIA, IRVINE Computational Models of Virus Dynamics DISSERTATION submitted in partial satisfaction of the requirements for the degree of DOCTOR OF PHILOSOPHY in Ecology and Evolutionary Biology by Sarah Marie Roy Dissertation Committee: Professor Dominik Wodarz, Chair Associate Professor Robin Bush Associate Professor Kevin Thornton 2015 © 2015 Sarah Marie Roy DEDICATION To my parents and to Hans, in loving memory ii TABLE OF CONTENTS Page LIST OF FIGURES iv ACKNOWLEDGMENTS v CURRICULUM VITAE vi ABSTRACT OF THE DISSERTATION vii INTRODUCTION 1 CHAPTER 1: Infection of HIV-specific CD4 T helper cells and the clonal 13 composition of the response CHAPTER 2: Tissue architecture, feedback regulation, and resilience to 46 viral infection CHAPTER 3: An Agent-Based Model of HIV Coinfection 71 iii LIST OF FIGURES Page Figure 1.1 Outcomes of model (1) assuming a single helper cell cell clone 21 Figure 1.2 Outcomes of model (2) assuming two independently regulated 24 helper cell clones Figure 1.3 Outcomes of model (2) depending on a and b 26 Figure 1.4 Outcomes of model (2) depending on r1 and r2 27 Figure 1.5 Outcomes of model (4) depending on CTL parameters 33 Figure 2.1 Dependence of Sfrac and Dfrac on replication rate, b 53 Figure 2.2 Tissue architecture and resilience to infection according to 55 model (2) Figure 2.3 Uncontrolled growth in the context of negative feedback 59 according to model (2) Figure 2.4 Two different virus persistence equilibria in the stem cell 61 infection model Figure 2.5 Stem cell infection rate vs. -

Rational Engineering of HCV Vaccines for Humoral Immunity
viruses Review To Include or Occlude: Rational Engineering of HCV Vaccines for Humoral Immunity Felicia Schlotthauer 1,2,†, Joey McGregor 1,2,† and Heidi E Drummer 1,2,3,* 1 Viral Entry and Vaccines Group, Burnet Institute, Melbourne, VIC 3004, Australia; [email protected] (F.S.); [email protected] (J.M.) 2 Department of Microbiology and Immunology, Peter Doherty Institute for Infection and Immunity, University of Melbourne, Melbourne, VIC 3000, Australia 3 Department of Microbiology, Monash University, Clayton, VIC 3800, Australia * Correspondence: [email protected]; Tel.: +61-392-822-179 † These authors contributed equally to this work. Abstract: Direct-acting antiviral agents have proven highly effective at treating existing hepatitis C infections but despite their availability most countries will not reach the World Health Organization targets for elimination of HCV by 2030. A prophylactic vaccine remains a high priority. Whilst early vaccines focused largely on generating T cell immunity, attention is now aimed at vaccines that gen- erate humoral immunity, either alone or in combination with T cell-based vaccines. High-resolution structures of hepatitis C viral glycoproteins and their interaction with monoclonal antibodies isolated from both cleared and chronically infected people, together with advances in vaccine technologies, provide new avenues for vaccine development. Keywords: glycoprotein E2; vaccine development; humoral immunity Citation: Schlotthauer, F.; McGregor, J.; Drummer, H.E To Include or 1. Introduction Occlude: Rational Engineering of HCV Vaccines for Humoral Immunity. The development of direct-acting antiviral agents (DAA) with their ability to cure Viruses 2021, 13, 805. https:// infection in >95% of those treated was heralded as the key to eliminating hepatitis C globally. -

Dissecting Human Antibody Responses Against Influenza a Viruses and Antigenic Changes That Facilitate Immune Escape
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations 2018 Dissecting Human Antibody Responses Against Influenza A Viruses And Antigenic Changes That Facilitate Immune Escape Seth J. Zost University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Allergy and Immunology Commons, Immunology and Infectious Disease Commons, Medical Immunology Commons, and the Virology Commons Recommended Citation Zost, Seth J., "Dissecting Human Antibody Responses Against Influenza A Viruses And Antigenic Changes That Facilitate Immune Escape" (2018). Publicly Accessible Penn Dissertations. 3211. https://repository.upenn.edu/edissertations/3211 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/3211 For more information, please contact [email protected]. Dissecting Human Antibody Responses Against Influenza A Viruses And Antigenic Changes That Facilitate Immune Escape Abstract Influenza A viruses pose a serious threat to public health, and seasonal circulation of influenza viruses causes substantial morbidity and mortality. Influenza viruses continuously acquire substitutions in the surface glycoproteins hemagglutinin (HA) and neuraminidase (NA). These substitutions prevent the binding of pre-existing antibodies, allowing the virus to escape population immunity in a process known as antigenic drift. Due to antigenic drift, individuals can be repeatedly infected by antigenically distinct influenza strains over the course of their life. Antigenic drift undermines the effectiveness of our seasonal influenza accinesv and our vaccine strains must be updated on an annual basis due to antigenic changes. In order to understand antigenic drift it is essential to know the sites of antibody binding as well as the substitutions that facilitate viral escape from immunity. -

Research Directions After the First Hepatitis C Vaccine Efficacy Trial
viruses Review Where to Next? Research Directions after the First Hepatitis C Vaccine Efficacy Trial Christopher C. Phelps 1 , Christopher M. Walker 1,2 and Jonathan R. Honegger 1,2,*,† 1 Center for Vaccines and Immunity, Abigail Wexner Research Institute at Nationwide Children’s Hospital, Columbus, OH 43205, USA; [email protected] (C.C.P.); [email protected] (C.M.W.) 2 Department of Pediatrics, The Ohio State University College of Medicine, Columbus, OH 43210, USA * Correspondence: [email protected] † Current address: Abigail Wexner Research Institute at Nationwide Children’s Hospital, 700 Children’s Drive, WA4020, Columbus, OH 43205, USA. Abstract: Thirty years after its discovery, the hepatitis C virus (HCV) remains a leading cause of liver disease worldwide. Given that many countries continue to experience high rates of transmission despite the availability of potent antiviral therapies, an effective vaccine is seen as critical for the elimination of HCV. The recent failure of the first vaccine efficacy trial for the prevention of chronic HCV confirmed suspicions that this virus will be a challenging vaccine target. Here, we examine the published data from this first efficacy trial along with the earlier clinical and pre-clinical studies of the vaccine candidate and then discuss three key research directions expected to be important in ongoing and future HCV vaccine development. These include the following: 1. design of novel immunogens that generate immune responses to genetically diverse HCV genotypes and subtypes, 2. strategies to elicit broadly neutralizing antibodies against envelope glycoproteins in addition to cytotoxic and helper T cell responses, and 3. -

All Qatar Residents to Get Covid Vaccine Free
INDEX QATAR 2-6,12 COMMENT 10 BUSINESS | Page 1 QATAR | Page 12 ARAB WORLD 7 BUSINESS 1-12 Qatar private INTERNATIONAL 7-9,11 SPORTS 1-8 Last Covid-19 sector bounces DOW JONES QE NYMEX patient back on lift ing discharged 28,148.64 9,956.66 39.36 of Covid-19 +465.83 +3.15 +2.31 from Lebsear +1.68% +0.03% +6.23% curbs: QFC Latest Figures Field Hospital published in QATAR since 1978 TUESDAY Vol. XXXXI No. 11693 October 6, 2020 Safar 19, 1442 AH GULF TIMES www. gulf-times.com 2 Riyals PM offers condolences to Amir of Kuwait HE the Prime Minister and Minister of Interior Sheikh Khalid bin Khalifa bin Abdulaziz al-Thani yesterday off ered condolences to the Amir of Kuwait, Sheikh Nawaf al-Ahmad al-Jaber al-Sabah, on the death of Sheikh Sabah al-Ahmad al-Jaber al-Sabah, praying to Allah to have mercy on the soul of the deceased and to rest it in peace in Paradise. The prime minister also off ered condolences to members of the ruling family and ranking off icials. Ministers and members of the off icial delegation accompanying the prime minister also off ered their condolences. Two held for violating PM meets Afghan president home quarantine rules Competent authorities arrested yesterday two people who violated All Qatar residents to the requirements of the home quarantine, which they committed to following, in accordance with the procedures of the health authorities in the country. The get Covid vaccine free two persons being referred to prosecution are: Albert Mondano Oshavillo and Nasser Ghaidan By Ayman Adly is not yet clear whether the upcoming Mohamed al-Hatheeth al-Qahtani. -
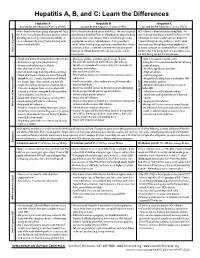
Hepatitis A, B, and C: Learn the Differences
Hepatitis A, B, and C: Learn the Differences Hepatitis A Hepatitis B Hepatitis C caused by the hepatitis A virus (HAV) caused by the hepatitis B virus (HBV) caused by the hepatitis C virus (HCV) HAV is found in the feces (poop) of people with hepa- HBV is found in blood and certain body fluids. The virus is spread HCV is found in blood and certain body fluids. The titis A and is usually spread by close personal contact when blood or body fluid from an infected person enters the body virus is spread when blood or body fluid from an HCV- (including sex or living in the same household). It of a person who is not immune. HBV is spread through having infected person enters another person’s body. HCV can also be spread by eating food or drinking water unprotected sex with an infected person, sharing needles or is spread through sharing needles or “works” when contaminated with HAV. “works” when shooting drugs, exposure to needlesticks or sharps shooting drugs, through exposure to needlesticks on the job, or from an infected mother to her baby during birth. or sharps on the job, or sometimes from an infected How is it spread? Exposure to infected blood in ANY situation can be a risk for mother to her baby during birth. It is possible to trans- transmission. mit HCV during sex, but it is not common. • People who wish to be protected from HAV infection • All infants, children, and teens ages 0 through 18 years There is no vaccine to prevent HCV. -

Understanding Human Astrovirus from Pathogenesis to Treatment
University of Tennessee Health Science Center UTHSC Digital Commons Theses and Dissertations (ETD) College of Graduate Health Sciences 6-2020 Understanding Human Astrovirus from Pathogenesis to Treatment Virginia Hargest University of Tennessee Health Science Center Follow this and additional works at: https://dc.uthsc.edu/dissertations Part of the Diseases Commons, Medical Sciences Commons, and the Viruses Commons Recommended Citation Hargest, Virginia (0000-0003-3883-1232), "Understanding Human Astrovirus from Pathogenesis to Treatment" (2020). Theses and Dissertations (ETD). Paper 523. http://dx.doi.org/10.21007/ etd.cghs.2020.0507. This Dissertation is brought to you for free and open access by the College of Graduate Health Sciences at UTHSC Digital Commons. It has been accepted for inclusion in Theses and Dissertations (ETD) by an authorized administrator of UTHSC Digital Commons. For more information, please contact [email protected]. Understanding Human Astrovirus from Pathogenesis to Treatment Abstract While human astroviruses (HAstV) were discovered nearly 45 years ago, these small positive-sense RNA viruses remain critically understudied. These studies provide fundamental new research on astrovirus pathogenesis and disruption of the gut epithelium by induction of epithelial-mesenchymal transition (EMT) following astrovirus infection. Here we characterize HAstV-induced EMT as an upregulation of SNAI1 and VIM with a down regulation of CDH1 and OCLN, loss of cell-cell junctions most notably at 18 hours post-infection (hpi), and loss of cellular polarity by 24 hpi. While active transforming growth factor- (TGF-) increases during HAstV infection, inhibition of TGF- signaling does not hinder EMT induction. However, HAstV-induced EMT does require active viral replication.