Insulin-Like Growth Factor Pathway and the Thyroid
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

HER2 Inhibition in Gastro-Oesophageal Cancer: a Review Drawing on Lessons Learned from Breast Cancer
Submit a Manuscript: http://www.f6publishing.com World J Gastrointest Oncol 2018 July 15; 10(7): 159-171 DOI: 10.4251/wjgo.v10.i7.159 ISSN 1948-5204 (online) REVIEW HER2 inhibition in gastro-oesophageal cancer: A review drawing on lessons learned from breast cancer Hazel Lote, Nicola Valeri, Ian Chau Hazel Lote, Nicola Valeri, Centre for Molecular Pathology, Accepted: May 30, 2018 Institute of Cancer Research, Sutton SM2 5NG, United Kingdom Article in press: May 30, 2018 Published online: July 15, 2018 Hazel Lote, Nicola Valeri, Ian Chau, Department of Medicine, Royal Marsden Hospital, Sutton SM2 5PT, United Kingdom ORCID number: Hazel Lote (0000-0003-1172-0372); Nicola Valeri (0000-0002-5426-5683); Ian Chau (0000-0003-0286-8703). Abstract Human epidermal growth factor receptor 2 (HER2)- Author contributions: Lote H wrote the original manuscript and revised it following peer review comments; Valeri N reviewed inhibition is an important therapeutic strategy in HER2- the manuscript; Chau I reviewed and contributed to the content of amplified gastro-oesophageal cancer (GOC). A significant the manuscript. proportion of GOC patients display HER2 amplification, yet HER2 inhibition in these patients has not displayed Supported by National Health Service funding to the National the success seen in HER2 amplified breast cancer. Mu- Institute for Health Research Biomedical Research Centre at ch of the current evidence surrounding HER2 has been the Royal Marsden NHS Foundation Trust and The Institute of obtained from studies in breast cancer, and we are only re- Cancer Research, No. A62, No. A100, No. A101 and No. A159; Cancer Research UK funding, No. -

Pharmacokinetics and Exposure-Response Relationship of Teprotumumab Frst 3 Months, Unless Determined to Be Medically Necessary
Clinical Pharmacokinetics https://doi.org/10.1007/s40262-021-01003-3 ORIGINAL RESEARCH ARTICLE Pharmacokinetics and Exposure‑Response Relationship of Teprotumumab, an Insulin‑Like Growth Factor‑1 Receptor‑Blocking Antibody, in Thyroid Eye Disease Yan Xin1 · Fengyan Xu2 · Yuying Gao2 · Nivedita Bhatt1 · Jason Chamberlain1 · Saba Sile1 · Suzy Hammel1 · Robert J. Holt1 · Srini Ramanathan1 Accepted: 10 February 2021 © The Author(s) 2021 Abstract Background and Objective Thyroid eye disease (TED) is characterized by infammation/expansion of orbital tissues, prop- tosis, and diplopia. Teprotumumab is the frst US Food and Drug Administration-approved therapy for TED, administered as an initial intravenous infusion of 10 mg/kg followed by 20 mg/kg every 3 weeks for an additional seven infusions. The objec- tive of this article is to discuss the pharmacokinetics and exposure-response profle for teprotumumab in patients with TED. Methods A population pharmacokinetic analysis was performed to characterize pharmacokinetics and select dosing in patients with TED. Exposure-response was evaluated for efcacy (proptosis response, clinical activity score categorical response, and diplopia response) and safety (hyperglycemia, muscle spasms, and hearing impairment) parameters. Results Teprotumumab pharmacokinetics was linear in patients with TED, with low systemic clearance (0.334 L/day), low volume of distribution (3.9 and 4.2 L for the central and peripheral compartment, respectively), and a long elimination half- life (19.9 days). The approved dosing regimen provided > 20 µg/mL for > 90% insulin-like growth factor 1 receptor saturation throughout the dosing interval. Model-predicted mean (± standard deviation) steady-state area under the concentration-time curve, peak, and trough concentrations in patients with TED were 131 (± 30.9) mg∙h/mL, 643 (± 130) µg/mL, and 157 (± 50.6) µg/mL, respectively. -

Or Ramucirumab (IMC-1121B) Plus Mitoxantrone and Prednisone in Men with Metastatic Docetaxel-Pretreated Castration-Resistant Prostate Cancer
European Journal of Cancer (2015) 51, 1714– 1724 Available at www.sciencedirect.com ScienceDirect journal homepage: www.ejcancer.com A randomised non-comparative phase II trial of cixutumumab (IMC-A12) or ramucirumab (IMC-1121B) plus mitoxantrone and prednisone in men with metastatic docetaxel-pretreated castration-resistant prostate cancer Maha Hussain a,1,⇑, Dana Rathkopf b,1, Glenn Liu c,1, Andrew Armstrong d,1, Wm. Kevin Kelly e, Anna Ferrari f, John Hainsworth g, Adarsh Joshi h, Rebecca R. Hozak i, Ling Yang h, Jonathan D. Schwartz h,2, Celestia S. Higano j,1 a University of Michigan Comprehensive Cancer Center, Ann Arbor, MI, United States b Memorial Sloan-Kettering, New York, NY, United States c University of Wisconsin, Carbone Cancer Center, Madison, WI, United States d Duke Cancer Institute and Duke Prostate Center, Duke University, Durham, NC, United States e Thomas Jefferson University, Philadelphia, PA, United States f New York University Clinical Cancer Center, New York, NY, United States g Sarah Cannon Research Institute, Nashville, TN, United States h Eli Lilly and Company, Bridgewater, NJ, United States i Eli Lilly and Company, Indianapolis, IN, United States j University of Washington, Fred Hutchinson Cancer Research Center, Seattle, WA, United States Received 11 February 2015; received in revised form 27 April 2015; accepted 10 May 2015 Available online 13 June 2015 KEYWORDS Abstract Background: Cixutumumab, a human monoclonal antibody (HuMAb), targets the Ramucirumab insulin-like growth factor receptor. Ramucirumab is a recombinant HuMAb that binds to vas- Cixutumumab cular endothelial growth factor receptor-2. A non-comparative randomised phase II study Mitoxantrone evaluated cixutumumab or ramucirumab plus mitoxantrone and prednisone (MP) in Prednisone metastatic castration-resistant prostate cancer (mCRPC). -

Oregon Medicaid Pharmaceutical Services Prior Authorization Criteria
Oregon Medicaid Pharmaceutical Services Prior Authorization Criteria HEALTH SYSTEMS DIVISION Prior authorization (PA) criteria for fee-for-service prescriptions for Oregon Health Plan clients March 1, 2021 Contents Contents ................................................................................................................................................................ 2 Introduction........................................................................................................................................................... 7 About this guide ......................................................................................................................................... 7 How to use this guide ................................................................................................................................. 7 Administrative rules and supplemental information .................................................................................. 7 Update information............................................................................................................................................... 8 Effective March 1, 2021 ............................................................................................................................ 8 Substantive updates and new criteria ............................................................................................. 8 Clerical changes ............................................................................................................................ -

IGF System in Sarcomas: a Crucial Pathway with Many Unknowns to Exploit for Therapy
61 1 Journal of Molecular C Mancarella and Targeting IGF system in sarcoma 61:1 T45–T60 Endocrinology K Scotlandi THEMATIC REVIEW 40 YEARS OF IGF1 IGF system in sarcomas: a crucial pathway with many unknowns to exploit for therapy Caterina Mancarella and Katia Scotlandi Experimental Oncology Lab, CRS Development of Biomolecular Therapies, Orthopaedic Rizzoli Institute, Bologna, Italy Correspondence should be addressed to K Scotlandi: [email protected] This paper forms part of a special section on 40 Years of IGF1. The guest editors for this section were Derek LeRoith and Emily Gallagher. Abstract The insulin-like growth factor (IGF) system has gained substantial interest due to its Key Words involvement in regulating cell proliferation, differentiation and survival during anoikis f sarcomas and after conventional and targeted therapies. However, results from clinical trials have f IGF system been largely disappointing, with only a few but notable exceptions, such as trials targeting f targeted therapy sarcomas, especially Ewing sarcoma. This review highlights key studies focusing on IGF f clinical trials signaling in sarcomas, specifically studies underscoring the properties that make this system an attractive therapeutic target and identifies new relationships that may be exploited. This review discusses the potential roles of IGF2 mRNA-binding proteins (IGF2BPs), discoidin domain receptors (DDRs) and metalloproteinase pregnancy-associated plasma protein-A (PAPP-A) in regulating the IGF system. Deeper investigation of these novel regulators of the IGF system may help us to further elucidate the spatial and temporal control of the IGF Journal of Molecular axis, as understanding the control of this axis is essential for future clinical studies. -

Interim Report for the First Quarter of 2020
Genmab Announces Financial Results for the First Quarter of 2020 May 6, 2020; Copenhagen, Denmark; Interim Report for the First Quarter Ended March 31, 2020 Highlights DARZALEX® (daratumumab) net sales increased approximately 49% compared to the first quarter of 2019 to USD 937 million, resulting in royalty income of DKK 775 million DARZALEX approved in Europe in combination with bortezomib, thalidomide and dexamethasone for the treatment of adult patients with newly diagnosed multiple myeloma who are eligible for autologous stem cell transplant U.S. FDA approved TEPEZZA™ (teprotumumab-trbw), developed and commercialized by Horizon Therapeutics, for thyroid eye disease U.S. FDA accepted, with priority review, Novartis’ supplemental Biologics License Application for subcutaneous ofatumumab in relapsing multiple sclerosis Anthony Pagano appointed Chief Financial Officer Anthony Mancini appointed Chief Operating Officer “Despite the unprecedented challenges posed by the coronavirus (COVID-19) pandemic, we will continue to invest in our innovative proprietary products, technologies and capabilities and use our world-class expertise in antibody drug development to create truly differentiated products with the potential to help cancer patients. While Genmab is closely monitoring the developments in the rapidly evolving landscape, we are extremely fortunate to have a solid financial foundation and a fabulous and committed team to carry us through these uncertain times,” said Jan van de Winkel, Ph.D., Chief Executive Officer of Genmab. Financial Performance First Quarter of 2020 Revenue was DKK 892 million in the first quarter of 2020 compared to DKK 591 million in the first quarter of 2019. The increase of DKK 301 million, or 51%, was mainly driven by higher DARZALEX royalties. -

Therapeutic Targeting of the IGF Axis
cells Review Therapeutic Targeting of the IGF Axis Eliot Osher and Valentine M. Macaulay * Department of Oncology, University of Oxford, Oxford, OX3 7DQ, UK * Correspondence: [email protected]; Tel.: +44-1865617337 Received: 8 July 2019; Accepted: 9 August 2019; Published: 14 August 2019 Abstract: The insulin like growth factor (IGF) axis plays a fundamental role in normal growth and development, and when deregulated makes an important contribution to disease. Here, we review the functions mediated by ligand-induced IGF axis activation, and discuss the evidence for the involvement of IGF signaling in the pathogenesis of cancer, endocrine disorders including acromegaly, diabetes and thyroid eye disease, skin diseases such as acne and psoriasis, and the frailty that accompanies aging. We discuss the use of IGF axis inhibitors, focusing on the different approaches that have been taken to develop effective and tolerable ways to block this important signaling pathway. We outline the advantages and disadvantages of each approach, and discuss progress in evaluating these agents, including factors that contributed to the failure of many of these novel therapeutics in early phase cancer trials. Finally, we summarize grounds for cautious optimism for ongoing and future studies of IGF blockade in cancer and non-malignant disorders including thyroid eye disease and aging. Keywords: IGF; type 1 IGF receptor; IGF-1R; cancer; acromegaly; ophthalmopathy; IGF inhibitor 1. Introduction Insulin like growth factors (IGFs) are small (~7.5 kDa) ligands that play a critical role in many biological processes including proliferation and protection from apoptosis and normal somatic growth and development [1]. IGFs are members of a ligand family that includes insulin, a dipeptide comprised of A and B chains linked via two disulfide bonds, with a third disulfide linkage within the A chain. -

Primary and Acquired Resistance to Immunotherapy in Lung Cancer: Unveiling the Mechanisms Underlying of Immune Checkpoint Blockade Therapy
cancers Review Primary and Acquired Resistance to Immunotherapy in Lung Cancer: Unveiling the Mechanisms Underlying of Immune Checkpoint Blockade Therapy Laura Boyero 1 , Amparo Sánchez-Gastaldo 2, Miriam Alonso 2, 1 1,2,3, , 1,2, , José Francisco Noguera-Uclés , Sonia Molina-Pinelo * y and Reyes Bernabé-Caro * y 1 Institute of Biomedicine of Seville (IBiS) (HUVR, CSIC, Universidad de Sevilla), 41013 Seville, Spain; [email protected] (L.B.); [email protected] (J.F.N.-U.) 2 Medical Oncology Department, Hospital Universitario Virgen del Rocio, 41013 Seville, Spain; [email protected] (A.S.-G.); [email protected] (M.A.) 3 Centro de Investigación Biomédica en Red de Cáncer (CIBERONC), 28029 Madrid, Spain * Correspondence: [email protected] (S.M.-P.); [email protected] (R.B.-C.) These authors contributed equally to this work. y Received: 16 November 2020; Accepted: 9 December 2020; Published: 11 December 2020 Simple Summary: Immuno-oncology has redefined the treatment of lung cancer, with the ultimate goal being the reactivation of the anti-tumor immune response. This has led to the development of several therapeutic strategies focused in this direction. However, a high percentage of lung cancer patients do not respond to these therapies or their responses are transient. Here, we summarized the impact of immunotherapy on lung cancer patients in the latest clinical trials conducted on this disease. As well as the mechanisms of primary and acquired resistance to immunotherapy in this disease. Abstract: After several decades without maintained responses or long-term survival of patients with lung cancer, novel therapies have emerged as a hopeful milestone in this research field. -

Horizon Therapeutics Secures US FDA Approval For
Horizon Therapeutics Secures U.S. FDA Approval for Teprotumumab, for the Treatment of Thyroid Eye Disease (TED) Media Release Copenhagen, Denmark, January 23, 2020 • Third Genmab-created product approved by the U.S. FDA Genmab A/S (Nasdaq: GMAB) announced today that the U.S. Food and Drug Administration (U.S. FDA) has granted approval to Horizon Therapeutics (Nasdaq: HZNP) for the use of teprotumumab, under the trade name TEPEZZA™ (teprotumumab-trbw), for the treatment of Thyroid Eye Disease (TED). TEPEZZA, the first and only U.S. FDA-approved medicine for the treatment of TED, was developed by and is manufactured by Horizon, completing a long development program that began when Genmab created the molecule nearly two decades ago. Horizon submitted the Biologics License Application for teprotumumab, which received Priority Review, Orphan Drug, Fast Track and Breakthrough Therapy designations from the FDA. Teprotumumab was created by Genmab under a collaboration with Roche and development of the product is now being conducted by Horizon under a license from Roche. Under the terms of Genmab’s agreement with Roche, Genmab will receive mid- single digit royalties on sales of TEPEZZA. “We would like to congratulate Horizon on this exciting approval, and we are very pleased that patients in the United States living with thyroid eye disease, a vision-threatening autoimmune condition, will now have this important new treatment option,” said Jan van de Winkel, Ph.D., Chief Executive Officer of Genmab. “We also believe that this approval, the third for a Genmab-created product following Arzerra® and DARZALEX®, reflects our company’s proven track record in improving the lives of patients by creating breakthrough innovative antibody products.” For more information about TEPEZZA and TED, please see the press release issued by Horizon Therapeutics. -
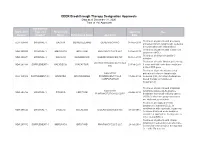
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

The Two Tontti Tudiul Lui Hi Ha Unit
THETWO TONTTI USTUDIUL 20170267753A1 LUI HI HA UNIT ( 19) United States (12 ) Patent Application Publication (10 ) Pub. No. : US 2017 /0267753 A1 Ehrenpreis (43 ) Pub . Date : Sep . 21 , 2017 ( 54 ) COMBINATION THERAPY FOR (52 ) U .S . CI. CO - ADMINISTRATION OF MONOCLONAL CPC .. .. CO7K 16 / 241 ( 2013 .01 ) ; A61K 39 / 3955 ANTIBODIES ( 2013 .01 ) ; A61K 31 /4706 ( 2013 .01 ) ; A61K 31 / 165 ( 2013 .01 ) ; CO7K 2317 /21 (2013 . 01 ) ; (71 ) Applicant: Eli D Ehrenpreis , Skokie , IL (US ) CO7K 2317/ 24 ( 2013. 01 ) ; A61K 2039/ 505 ( 2013 .01 ) (72 ) Inventor : Eli D Ehrenpreis, Skokie , IL (US ) (57 ) ABSTRACT Disclosed are methods for enhancing the efficacy of mono (21 ) Appl. No. : 15 /605 ,212 clonal antibody therapy , which entails co - administering a therapeutic monoclonal antibody , or a functional fragment (22 ) Filed : May 25 , 2017 thereof, and an effective amount of colchicine or hydroxy chloroquine , or a combination thereof, to a patient in need Related U . S . Application Data thereof . Also disclosed are methods of prolonging or increasing the time a monoclonal antibody remains in the (63 ) Continuation - in - part of application No . 14 / 947 , 193 , circulation of a patient, which entails co - administering a filed on Nov. 20 , 2015 . therapeutic monoclonal antibody , or a functional fragment ( 60 ) Provisional application No . 62/ 082, 682 , filed on Nov . of the monoclonal antibody , and an effective amount of 21 , 2014 . colchicine or hydroxychloroquine , or a combination thereof, to a patient in need thereof, wherein the time themonoclonal antibody remains in the circulation ( e . g . , blood serum ) of the Publication Classification patient is increased relative to the same regimen of admin (51 ) Int . -

(12) Patent Application Publication (10) Pub. No.: US 2017/0172932 A1 Peyman (43) Pub
US 20170172932A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2017/0172932 A1 Peyman (43) Pub. Date: Jun. 22, 2017 (54) EARLY CANCER DETECTION AND A 6LX 39/395 (2006.01) ENHANCED IMMUNOTHERAPY A61R 4I/00 (2006.01) (52) U.S. Cl. (71) Applicant: Gholam A. Peyman, Sun City, AZ CPC .......... A61K 9/50 (2013.01); A61K 39/39558 (US) (2013.01); A61K 4I/0052 (2013.01); A61 K 48/00 (2013.01); A61K 35/17 (2013.01); A61 K (72) Inventor: sham A. Peyman, Sun City, AZ 35/15 (2013.01); A61K 2035/124 (2013.01) (21) Appl. No.: 15/143,981 (57) ABSTRACT (22) Filed: May 2, 2016 A method of therapy for a tumor or other pathology by administering a combination of thermotherapy and immu Related U.S. Application Data notherapy optionally combined with gene delivery. The combination therapy beneficially treats the tumor and pre (63) Continuation-in-part of application No. 14/976,321, vents tumor recurrence, either locally or at a different site, by filed on Dec. 21, 2015. boosting the patient’s immune response both at the time or original therapy and/or for later therapy. With respect to Publication Classification gene delivery, the inventive method may be used in cancer (51) Int. Cl. therapy, but is not limited to such use; it will be appreciated A 6LX 9/50 (2006.01) that the inventive method may be used for gene delivery in A6 IK 35/5 (2006.01) general. The controlled and precise application of thermal A6 IK 4.8/00 (2006.01) energy enhances gene transfer to any cell, whether the cell A 6LX 35/7 (2006.01) is a neoplastic cell, a pre-neoplastic cell, or a normal cell.