Spinal Reflexes & Neuronal Integration
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neural Control of Movement: Motor Neuron Subtypes, Proprioception and Recurrent Inhibition
List of Papers This thesis is based on the following papers, which are referred to in the text by their Roman numerals. I Enjin A, Rabe N, Nakanishi ST, Vallstedt A, Gezelius H, Mem- ic F, Lind M, Hjalt T, Tourtellotte WG, Bruder C, Eichele G, Whelan PJ, Kullander K (2010) Identification of novel spinal cholinergic genetic subtypes disclose Chodl and Pitx2 as mark- ers for fast motor neurons and partition cells. J Comp Neurol 518:2284-2304. II Wootz H, Enjin A, Wallen-Mackenzie Å, Lindholm D, Kul- lander K (2010) Reduced VGLUT2 expression increases motor neuron viability in Sod1G93A mice. Neurobiol Dis 37:58-66 III Enjin A, Leao KE, Mikulovic S, Le Merre P, Tourtellotte WG, Kullander K. 5-ht1d marks gamma motor neurons and regulates development of sensorimotor connections Manuscript IV Enjin A, Leao KE, Eriksson A, Larhammar M, Gezelius H, Lamotte d’Incamps B, Nagaraja C, Kullander K. Development of spinal motor circuits in the absence of VIAAT-mediated Renshaw cell signaling Manuscript Reprints were made with permission from the respective publishers. Cover illustration Carousel by Sasha Svensson Contents Introduction.....................................................................................................9 Background...................................................................................................11 Neural control of movement.....................................................................11 The motor neuron.....................................................................................12 Organization -

GDE2 Regulates Subtype-Specific Motor Neuron Generation Through
Neuron Article GDE2 Regulates Subtype-Specific Motor Neuron Generation through Inhibition of Notch Signaling Priyanka Sabharwal,1 Changhee Lee,1 Sungjin Park,1 Meenakshi Rao,1,2 and Shanthini Sockanathan1,* 1The Solomon Snyder Department of Neuroscience, The Johns Hopkins University School of Medicine, PCTB1004, 725 N. Wolfe Street, Baltimore, MD 21205, USA 2Present address: Department of Pediatrics, Children’s Hospital Boston, 300 Longwood Avenue, Boston, MA 02115, USA *Correspondence: [email protected] DOI 10.1016/j.neuron.2011.07.028 SUMMARY which is innervated by specific groups of motor neurons. Indi- vidual motor neuron groups are highly organized in terms of their The specification of spinal interneuron and motor cell body distribution, projection patterns, and function and neuron identities initiates within progenitor cells, consist of force-generating alpha motor neurons that innervate while motor neuron subtype diversification is regu- extrafusal muscle fibers and stretch-sensitive gamma motor lated by hierarchical transcriptional programs imple- neurons that innervate intrafusal muscle fibers of the muscle mented postmitotically. Here we find that mice lack- spindles (Dasen and Jessell, 2009; reviewed in Kanning et al., ing GDE2, a six-transmembrane protein that triggers 2010). The integration of input from both alpha and gamma motor neurons is essential for coordinated motor movement to motor neuron generation, exhibit selective losses occur (Kanning et al., 2010). of distinct motor neuron subtypes, specifically in How is diversity engendered in developing motor neurons? defined subsets of limb-innervating motor pools All motor neurons initially derive from ventral progenitor cells that correlate with the loss of force-generating alpha that are specified to become Olig2+ motor neuron progenitors motor neurons. -

Reduction of Ephrin-A5 Aggravates Disease Progression in Amyotrophic
Rué et al. Acta Neuropathologica Communications (2019) 7:114 https://doi.org/10.1186/s40478-019-0759-6 RESEARCH Open Access Reduction of ephrin-A5 aggravates disease progression in amyotrophic lateral sclerosis Laura Rué1,2 , Patrick Oeckl3, Mieke Timmers1,2, Annette Lenaerts1,2, Jasmijn van der Vos1,2, Silke Smolders1,2, Lindsay Poppe1,2, Antina de Boer1,2, Ludo Van Den Bosch1,2, Philip Van Damme1,2,4, Jochen H. Weishaupt3, Albert C. Ludolph3, Markus Otto3, Wim Robberecht1,4 and Robin Lemmens1,2,4* Abstract Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease that affects motor neurons in the brainstem, spinal cord and motor cortex. ALS is characterized by genetic and clinical heterogeneity, suggesting the existence of genetic factors that modify the phenotypic expression of the disease. We previously identified the axonal guidance EphA4 receptor, member of the Eph-ephrin system, as an ALS disease-modifying factor. EphA4 genetic inhibition rescued the motor neuron phenotype in zebrafish and a rodent model of ALS. Preventing ligands from binding to the EphA4 receptor also successfully improved disease, suggesting a role for EphA4 ligands in ALS. One particular ligand, ephrin-A5, is upregulated in reactive astrocytes after acute neuronal injury and inhibits axonal regeneration. Moreover, it plays a role during development in the correct pathfinding of motor axons towards their target limb muscles. We hypothesized that a constitutive reduction of ephrin-A5 signalling would benefit disease progression in a rodent model for ALS. We discovered that in the spinal cord of control and symptomatic ALS mice ephrin-A5 was predominantly expressed in neurons. -

Cortex Brainstem Spinal Cord Thalamus Cerebellum Basal Ganglia
Harvard-MIT Division of Health Sciences and Technology HST.131: Introduction to Neuroscience Course Director: Dr. David Corey Motor Systems I 1 Emad Eskandar, MD Motor Systems I - Muscles & Spinal Cord Introduction Normal motor function requires the coordination of multiple inter-elated areas of the CNS. Understanding the contributions of these areas to generating movements and the disturbances that arise from their pathology are important challenges for the clinician and the scientist. Despite the importance of diseases that cause disorders of movement, the precise function of many of these areas is not completely clear. The main constituents of the motor system are the cortex, basal ganglia, cerebellum, brainstem, and spinal cord. Cortex Basal Ganglia Cerebellum Thalamus Brainstem Spinal Cord In very broad terms, cortical motor areas initiate voluntary movements. The cortex projects to the spinal cord directly, through the corticospinal tract - also known as the pyramidal tract, or indirectly through relay areas in the brain stem. The cortical output is modified by two parallel but separate re entrant side loops. One loop involves the basal ganglia while the other loop involves the cerebellum. The final outputs for the entire system are the alpha motor neurons of the spinal cord, also called the Lower Motor Neurons. Cortex: Planning and initiation of voluntary movements and integration of inputs from other brain areas. Basal Ganglia: Enforcement of desired movements and suppression of undesired movements. Cerebellum: Timing and precision of fine movements, adjusting ongoing movements, motor learning of skilled tasks Brain Stem: Control of balance and posture, coordination of head, neck and eye movements, motor outflow of cranial nerves Spinal Cord: Spontaneous reflexes, rhythmic movements, motor outflow to body. -

Gamma Motor Neurons Survive and Exacerbate Alpha Motor Neuron Degeneration In
Gamma motor neurons survive and exacerbate alpha PNAS PLUS motor neuron degeneration in ALS Melanie Lalancette-Heberta,b, Aarti Sharmaa,b, Alexander K. Lyashchenkoa,b, and Neil A. Shneidera,b,1 aCenter for Motor Neuron Biology and Disease, Columbia University, New York, NY 10032; and bDepartment of Neurology, Columbia University, New York, NY 10032 Edited by Rob Brownstone, University College London, London, United Kingdom, and accepted by Editorial Board Member Fred H. Gage October 27, 2016 (received for review April 4, 2016) The molecular and cellular basis of selective motor neuron (MN) the muscle spindle and control the sensitivity of spindle afferent vulnerability in amyotrophic lateral sclerosis (ALS) is not known. In discharge (15); beta (β) skeletofusimotor neurons innervate both genetically distinct mouse models of familial ALS expressing intra- and extrafusal muscle (16). In addition to morphological mutant superoxide dismutase-1 (SOD1), TAR DNA-binding protein differences, distinct muscle targets, and the absence of primary 43 (TDP-43), and fused in sarcoma (FUS), we demonstrate selective afferent (IA)inputsonγ-MNs, these functional MN subtypes also degeneration of alpha MNs (α-MNs) and complete sparing of differ in their trophic requirements, and γ-MNs express high levels gamma MNs (γ-MNs), which selectively innervate muscle spindles. of the glial cell line-derived neurotropic factor (GDNF) receptor Resistant γ-MNs are distinct from vulnerable α-MNs in that they Gfrα1 (17). γ-MNs are also molecularly distinguished by the ex- lack synaptic contacts from primary afferent (IA) fibers. Elimination pression of other selective markers including the transcription α of these synapses protects -MNs in the SOD1 mutant, implicating factor Err3 (18), Wnt7A (19), the serotonin receptor 1d (5-ht1d) this excitatory input in MN degeneration. -
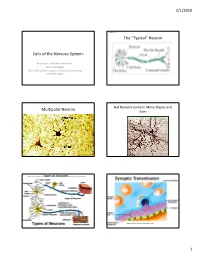
Cells of the Nervous System: the “Typical” Neuron Multipolar Neuron
2/1/2010 Book Fig. 1.1 The “Typical” Neuron Cells of the Nervous System: Neurons: cells that receive & send messages Glia: cells which support neuron functioning in many ways But Neurons Come in Many Shapes and Multipolar Neuron Sizes Types of Neurons Book Fig 1.1 Sensory Neuron Motor Neuron Some proteins serve as receptor sites. 1 2/1/2010 Best Known Neurotransmitters (study handout linked to syllabus) • Acetylcholine (ACh) • Norepinephrine (NE) • Dopamine (DA) • Serotonin or 5-Hydroxytryptamine (5HT) • GABA Released neurotransmitter must bind to specially shaped receptors like a key fitting into a lock. We now know there are multiple subtypes of receptors for each • Glutamate-most widespread excitatory neurotransmitter. transmitter Then the transmitter must be removed from the synapse either by reuptake or enzymatic breakdown. Here’s some background on ACh before we cover an Best Known Neurotransmitters Example of a neurotransmitter related disorder (FYI only – not completely up-to-date list of the number Acetylcholine (ACh) of identified receptor subtypes) • Acetylcholine (ACh) (7 receptor subtypes) • neurons using ACh are known as “cholinergic neurons”. • Examples: Norepinephrine (NE) (11 receptor subtypes) • motor neurons • Dopamine DA) (5 receptor subtypes) • parasympathetic neurons • many CNS neurons (in cortex, basal ganglia, hippocampus, • Serotonin (5HT) (14 receptor subtypes) brainstem) • GABA (2 receptor subtypes) • Different ACh receptor types on muscle (nicotinic) than in the nervous system (muscarinic) • Glutamate (10 receptor -

Mitigation of ALS Pathology by Neuron-Specific Inhibition of Nuclear Factor Kappa B Signaling
The Journal of Neuroscience, June 24, 2020 • 40(26):5137–5154 • 5137 Neurobiology of Disease Mitigation of ALS Pathology by Neuron-Specific Inhibition of Nuclear Factor Kappa B Signaling Kallol Dutta,1 Sai Sampath Thammisetty,1 Hejer Boutej,1 Christine Bareil,1 and Jean-Pierre Julien1,2 1CERVO Brain Research Centre, Québec City, Québec G1J 2G3, Canada, and 2Department of Psychiatry and Neuroscience, Université Laval, Québec City, Québec G1V 0A6, Canada To investigate the role of neuronal NF-jB activity in pathogenesis of amyotrophic lateral sclerosis (ALS), we generated trans- genic mice with neuron-specific expression of a super-repressor form of the NF-jB inhibitor (IjBa-SR), which were then crossed with mice of both sexes, expressing ALS-linked gene mutants for TAR DNA-binding protein (TDP-43) and superoxide dismutase 1 (SOD1). Remarkably, neuronal expression of IjBa-SR transgene in mice expressing TDP-43A315T or TDP-43G348C mice led to a decrease in cytoplasmic to nuclear ratio of human TDP-43. The mitigation of TDP-43 neuropathology by IjBa-SR, which is likely due to an induction of autophagy, was associated with amelioration of cognitive and motor deficits as well as reduc- tion of motor neuron loss and gliosis. Neuronal suppression of NF-jB activity in SOD1G93A mice also resulted in neuroprotection with reduction of misfolded SOD1 levels and significant extension of life span. The results suggest that neuronal NF-jB signaling constitutes a novel therapeutic target for ALS disease and related disorders with TDP-43 proteinopathy. Key words: amyotrophic lateral sclerosis; frontotemporal dementia; IjB suppressor; NF-jB; superoxide dismutase; TDP-43 Significance Statement This study reports that neuron-specific expression of IkB super-repressor mitigated behavioral and pathologic changes in transgenic mouse models of amyotrophic lateral sclerosis expressing mutant forms of either Tar DNA-binding protein 43 or superoxide dismutase. -

NERVOUS SYSTEM : LEC / 8 Physiology of Nerves and Muscles the Nervous System Is One of the Most Complicated System of the Body in Both Structure and Function
Medical physiology Lecturer Second Stage Hiba Hazim Saleh NERVOUS SYSTEM : LEC / 8 Physiology of nerves and muscles The nervous system is one of the most complicated system of the body in both structure and function. It senses physical and chemical changes in the internal and external environment, processes them, and then responds to maintain homeostasis. Voluntary activities, such as walking and talking, and involuntary activities, such as digestion and circulation, are coordinates, regulated, and integrated by the nervous system. The entire neural network of the body relies on the transmission of nervous impulses. Nervous impulses are electrochemical stimuli that travel from cell to cell as they send information from one area of the body to another. The speed at which this occurs is almost instantaneous, thus providing an immediate response change. (Figure -1. ) Muscles Muscle tissue is composed of contractile cells or fibers that provide movement of an organ or body part. Muscles contribute to posture, produce body heat, and act as a protective covering for internal organs. Muscles make up the bulk of the body. They have the ability to be excited by a stimulus, contract, relax, and return to their original size and shape. Whether muscles are attached to bones or to internal organs and blood vessels, their primary responsibility is movement. Apparent motion provided by muscles include walking and talking. Less apparent motion include the passage and elimination of food through the digestive system, propulsion of blood through the arteries, and contraction of the bladder to eliminate urine. (Figure -2 .) Figure -1 Figure-2 Never Cell (Neuron): Nerve Cell: Is a basic unit of nervous system. -

Nerve Cell Impulses
• Localization of Certain Neurons Neurotransmitters Nerve Conduction by: Mary V. Andrianopoulos, Ph.D Clarification: Types of Neuron • There may be none, one, or many dendrites composing part of a neuron. • No dendrite = a unipolar neuron • One dendrite = bipolar neuron • More than one dendrite = multipolar neuron. Multipolar neuron Bipolar neuron Unipolar neuron Localization of Neuron types • Unipolar: – found in most of body's sensory neurons – dendrites are the exposed branches connected to receptors – axon carries the action potential in to the CNS – Examples: posterior root ganglia + cranial nerves – Usually: have peripheral + central connections Localization of Neuron types • Bipolar: – retina, sensory cochlear, vestibular ganglion • Multipolar: (fibers) brain + spinal cord – found as motor neurons and interneurons – neuronal tractsÆ CNS – peripheral nervesÆ PNS Size of Neurons + their localization • Golgi I: – Fiber tracts: brain + spinal cord (PNS + motor) – (i.e., Pyramidal tract + Purkinje cells) • Golgi II: – Cerebral + cerebellar cortex – Often inhibitory – Out number Golgi I – Star-shaped appearance 2° short dendrites Histology of the Nervous System A review of Cell types 1) Neurons - the functional cells of the nervous system 2) Neuroglia (glial cells) - Long described as supporting cells of the nervous system, there is also a functional interdependence of neuroglial cells and neurons a) astrocytes - anchor neurons to blood vessels, regulate the micro-environment of neurons, and regulate transport of nutrients and wastes to and from neurons b) microglia- are phagocytic to defend against pathogens and monitor the condition of neurons c) ependymal - line the fluid-filled cavities of the brain and spinal column and play a role in production, transport, and circulation of the CSF. -

Control of Body Movement
Control of Body Movement Chapter 10 To learn how NS system and skeletal muscle interact with each other in the initiation and control of body movements 成大生理所 張雅雯老師 Motor control hierarchy 10.1 Motor control hierarchy運動控制之層次結構 Higher-center “command” neurons Generate an intention The middle-level structures Receive input and create/adjust motor program Local-level neurons Carry out the desired action Middle level of the motor control hierarchy Five components of the middle level of the motor control hierarchy Proprioception本體感覺 Afferent information about the position of the body and its parts in space Include balance, coordination and agility Enable the body to move and control limbs without directly looking at them With/without ever reaching the level of conscious perceptionhttps://draxe.com/wp-content/uploads/2016/01/ProprioceptionGraphic.jpg' http://www.4dancers.org/wp-content/uploads/2013/09/Picture-4.png Voluntary and involuntary actions Voluntary actions (1) the movement is accompanied by a conscious awareness of what we are doing and why we are doing it (2) our attention is directed toward the action or its purpose Involuntary actions Unconscious, automatic, reflex Almost all motor behavior involves both actions Not easy to make a distinction between these actions http://images.slideplayer.com/14/4207261/slides/slide_11.jpg; http://4.bp.blogspot.com/-KKp2zY_IeTc/UvoCbGoqs7I/AAAAAAAAA2Y/FM4DVIFL8ng/s1600/Slide09.jpg Local control of motor neurons 10.2 Motor neurons always form the final common pathway to the muscles Interneurons中間神經元 -

The Nervous System 1) Integration of Body Processes 2) Control of Voluntary Effectors (Skeletal Muscles), and Mediation of Voluntary Reflexes
1 © Jim Swan These slides are from class presentations, reformatted for static viewing. The content contained in these pages is also in the Class Notes pages in a narrative format. Best screen resolution for viewing is 1024 x 768. To change resolution click on start, then control panel, then display, then settings. If you are viewing this in Adobe Reader version 7 and are connected to the internet you will also be able to access the “enriched” links to notes and comments, as well as web pages including animations and videos. You will also be able to make your own notes and comments on the pages. Download the free reader from [Adobe.com] 1 Functions of the Nervous System 1) Integration of body processes 2) Control of voluntary effectors (skeletal muscles), and mediation of voluntary reflexes. 3) Control of involuntary effectors ( smooth muscle, cardiac muscle, glands) and mediation of autonomic reflexes (heart rate, blood pressure, glandular secretion, etc.) 4) Response to stimuli 5) Responsible for conscious thought and perception, emotions, personality, the mind. 2 These functions relate to control of the skeletal muscles discussed in Unit 2 as well as future discussion of reflexes, the brain, and the autonomic nervous system. 2 Structural Divisions of the Nervous System Central Nervous System (CNS) Brain Spinal Cord Peripheral Nervous System (PNS) nerves, ganglia, receptors 3 The central nervous system develops from the neural tube, while the peripheral nervous system develops from the neural crest cells. 3 Functional Divisions of the Nervous System 1) The Voluntary Nervous System - (a.k.a. somatic division) willful control of effectors (skeletal muscles), and conscious perception. -

Human Anatomy and Physiology I Laboratory
Human Anatomy and Physiology I Laboratory Histology of Nervous Tissue and The Spinal Cord This lab involves two laboratory exercises: 1) “Histology of Nervous Tissue”, and 2) “Spinal Cord, Spinal Nerves, and the Autonomic Nervous System”. Complete the Review Sheet for the entire first exercise, and the portion pertaining to the spinal cord for the second. The remainder of the second exercise will be completed in the next lab on the spinal and peripheral nerves. The quizzes are separate as well: for this lab take the quiz on nervous tissue and the spinal cord. Alternately, your instructor may have you turn in drawings of nerve tissue in lieu of the Review Sheets. Use the Virtual Microscope or other histology sites for images of nerve tissue. Click on the sound icon for the audio file (mp3 format) for each slide. There is also a link to a dowloadable mp4 video which can be played on an iPod. 1 Cells of the Nervous System: Glial Cells (neuroglia) and other supportive cells: These provide supportive functions for the nervous system. And new information stresses the functional interdependence of glial cells and neurons. Neurons: these are the functional cells of the nervous system, i.e. they conduct the impulses. 2 CNS Neuroglia a) Astrocytes - these cells anchor neurons to blood vessels, regulate the micro-environment of neurons, and regulate transport of nutrients and wastes to and from neurons. They are part of the blood-brain barrier. b) Microglia - these cells are phagocytic to defend against pathogens. They may also monitor the condition of neurons.