Impact of Ecotourism on Abundance, Diversity and Activity
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2019 Regional Leatherback Bycatch Prioritization Workshop
2019 Regional Leatherback Bycatch Prioritization Workshop * Attending both the Regional Bycatch Workshop attending the WIDECAST Annual Meeting (*) Attending only the Regional Bycatch Workshop NAME POSITION INSTITUTE ADDRESS PHONE EMAIL Canada: * Brianne Kelly Senior Specialist WWF-Canada 5251 Duke Street 902.482.1105 [email protected] Marine Ecosystems Duke Tower Suite ext. 3025 & Fisheries 1202 Halifax NS B3J 1P3 France/ French-Guiane * Dr. Damien Chevallier Researcher Centre National de 3 rue Michel-Ange +0612 97 10 54 [email protected] Recherche Délégation Alsace Scientifique – Inst 23 rue du Loess – Pluridisciplinaire BP20 Hubert Curien Strasbourg, (CNRS-IPHC) France * Nicolas Paranthoën Coordinateur du Plan Office National de la Kourou, Guyane +0694 13 77 44 [email protected] National d'Actions Chasse et de la française Tortues Marines en Faune Sauvage Guyane (ONCFS) * Rachel Berzins Cheffe de la cellule Office National de la Kourou, Guyane +0694 40 45 14 [email protected] technique de Guyane Chasse et de la française Faune Sauvage (ONCFS) 2019 Bycatch Prioritization Workshop, Paramaribo SURINAME 1 * Christelle Guyon Chargée de mission Direction de Rue Carlos +0594 29 68 60 Christelle.guyon@developpement- biodiversité marine l’Environnement, de Fineley CS 76003, durable.gouv.fr l’Aménagement et 97306 Cayenne du Logement Guyane française de Guyane * Nolwenn Cozannet Chargée du projet WWF France, 2, rue Gustave +0594 31 38 28 [email protected] Dauphin de Guyane bureau Guyane Charlery 97 300 CAYENNE -

Line Transect Surveys Underdetect Terrestrial Mammals: Implications for the Sustainability of Subsistence Hunting
RESEARCH ARTICLE Line Transect Surveys Underdetect Terrestrial Mammals: Implications for the Sustainability of Subsistence Hunting José M. V. Fragoso1☯¤*, Taal Levi2‡, Luiz F. B. Oliveira3‡, Jeffrey B. Luzar4‡, Han Overman5‡, Jane M. Read6‡, Kirsten M. Silvius7☯ 1 Stanford University, Stanford, CA, 94305–5020, United States of America, 2 Department of Fisheries and Wildlife, Oregon State University, Corvallis, OR, United States of America, 3 Departamento de Vertebrados, Museu Nacional, UFRJ, RJ, 20.940–040, Brazil, 4 Stanford University, Stanford, CA, 94305–5020, United States of America, 5 Environmental and Forest Biology, State University of New York-College of Environmental Science and Forestry, Syracuse, NY, 13210, United States of America, 6 Geography Department, Syracuse University, Syracuse, NY, 13244, United States of America, 7 Department of Forest Resources and Environmental Conservation, Virginia Tech, Blacksburg, VA, United States of America ☯ These authors contributed equally to this work. ¤ Current address: Biodiversity Science and Sustainability, California Academy of Sciences, San Francisco, California, United States of America OPEN ACCESS ‡ These authors contributed subsets of effort equally to this work. * [email protected] Citation: Fragoso JMV, Levi T, Oliveira LFB, Luzar JB, Overman H, Read JM, et al. (2016) Line Transect Surveys Underdetect Terrestrial Mammals: Implications for the Sustainability of Subsistence Abstract Hunting. PLoS ONE 11(4): e0152659. doi:10.1371/ journal.pone.0152659 Conservation of Neotropical game species must take into account the livelihood and food Editor: Mathew S. Crowther, University of Sydney, security needs of local human populations. Hunting management decisions should there- AUSTRALIA fore rely on abundance and distribution data that are as representative as possible of true Received: January 19, 2016 population sizes and dynamics. -

Dolichotis Patagonum (CAVIOMORPHA; CAVIIDAE; DOLICHOTINAE) Mastozoología Neotropical, Vol
Mastozoología Neotropical ISSN: 0327-9383 ISSN: 1666-0536 [email protected] Sociedad Argentina para el Estudio de los Mamíferos Argentina Silva Climaco das Chagas, Karine; Vassallo, Aldo I; Becerra, Federico; Echeverría, Alejandra; Fiuza de Castro Loguercio, Mariana; Rocha-Barbosa, Oscar LOCOMOTION IN THE FASTEST RODENT, THE MARA Dolichotis patagonum (CAVIOMORPHA; CAVIIDAE; DOLICHOTINAE) Mastozoología Neotropical, vol. 26, no. 1, 2019, -June, pp. 65-79 Sociedad Argentina para el Estudio de los Mamíferos Argentina Available in: https://www.redalyc.org/articulo.oa?id=45762554005 How to cite Complete issue Scientific Information System Redalyc More information about this article Network of Scientific Journals from Latin America and the Caribbean, Spain and Journal's webpage in redalyc.org Portugal Project academic non-profit, developed under the open access initiative Mastozoología Neotropical, 26(1):65-79, Mendoza, 2019 Copyright ©SAREM, 2019 Versión on-line ISSN 1666-0536 http://www.sarem.org.ar https://doi.org/10.31687/saremMN.19.26.1.0.06 http://www.sbmz.com.br Artículo LOCOMOTION IN THE FASTEST RODENT, THE MARA Dolichotis patagonum (CAVIOMORPHA; CAVIIDAE; DOLICHOTINAE) Karine Silva Climaco das Chagas1, 2, Aldo I. Vassallo3, Federico Becerra3, Alejandra Echeverría3, Mariana Fiuza de Castro Loguercio1 and Oscar Rocha-Barbosa1, 2 1 Laboratório de Zoologia de Vertebrados - Tetrapoda (LAZOVERTE), Departamento de Zoologia, IBRAG, Universidade do Estado do Rio de Janeiro, Maracanã, Rio de Janeiro, Brasil. 2 Programa de Pós-Graduação em Ecologia e Evolução do Instituto de Biologia/Uerj. 3 Laboratorio de Morfología Funcional y Comportamiento. Departamento de Biología; Instituto de Investigaciones Marinas y Costeras (CONICET); Universidad Nacional de Mar del Plata. -

Relative Rates of Molecular Evolution in Rodents and Their Symbionts
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1997 Relative Rates of Molecular Evolution in Rodents and Their yS mbionts. Theresa Ann Spradling Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Spradling, Theresa Ann, "Relative Rates of Molecular Evolution in Rodents and Their yS mbionts." (1997). LSU Historical Dissertations and Theses. 6527. https://digitalcommons.lsu.edu/gradschool_disstheses/6527 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type o f computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely afreet reproduction. hi the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. -

Special Update on Quito Process V Technical Meeting on Human
Special update on Quito Process AS OF NOVEMBER 2019 LATIN AMERICA AND THE CARIBBEAN AS OF NOVEMBER 2019 LATIN AMERICA AND THE CARIBBEAN V Technical Meeting on HumanV EMobilityNEZUVELEANNEZUE LofAN Venezuelan Citizens in the RegionREFUGEERSE F&U MGIEGERSA &N TMSIG INR ATNHTES R IENG TIOHNE REGION UNITED STATES UNITED STATES BOGOTA, COLOMBIA Havana\ MEXICO Havana\ MEXICO CUBA DOMINICAN 14-15 NOVEMBER 2019 CUBA DOMINICAN \ REPUBLIC Mexico \ HAITI \ PUERTO REPUBLIC Mexico JAMAICA \ Santo HAITI \ PUERTO City \BELIZE JAMAICA \ RICO City \ Domingo Santo RICO BELIZE Domingo \ HONDURAS CARIBBEAN SEA GUATEMALA \ \ HONDURAS CARIBBEAN SEA \ GUATEMALA \ EL SALVADOR NICARAGUA \ ARUBA CURACAO \ EL SALVADOR APoRrUt BA \NICARAGUA Spain CURACAO Port San José \ \ TRINIDAD Spain \ Panamá San JoséCaracas & TOBA\GO \ TRINIDAD PARTICIPATION COSTA \ \ Caracas Panamá & TOBAGO COSTA \ RICA PANAMA Georgetown RICA PANAMAVENEZUELA \ Georgetown VENEZ\UPaErLaAmaribo \ GUYANA Paramaribo \Bogotá \ CGayUeYnAneNA \ \Bogotá \ Cayenne COLOMBIA SURINAME COLOMBIA FRENCH SURINAME GUYANA FRENCH The fifth round of the Quito Process (Quito V – GUYANA \Quito \Quito ECUADOR Bogota Chapter) took place on 14-15 November ECUADOR (2019), with the participation of 12 States from PERU Latin America and the Caribbean, along with PERU \Lima BRAZIL key cooperating States, international financial \Lima BRAZIL \ BOLIVIA Brasilia \ BOLIVIA Brasilia institutions and other actors. \ \ Sucre Sucre PACIFIC PARAGUAY ATLANTIC PACIFIC PARAGUAY ATLANTIC OCEAN Asunción \ OCEAN The meeting follows up on four previous meetings OCEAN Asunción \ OCEAN QUITO V DECLARATION in September and November 2018, April and July QUITO V DECLARATION SIGNATORIES ARGENTINA SIGNATORIES ARGENTINA 2019, as part of a government-led initiative thatArge ntina \ URUGUAY Argentina \ URUGUAY Santiago \ \ Santiago \ Brazil Buenos Montevideo \ aims to harmonize the policies and practices of Brazil Aires Buenos Montevideo Chile CHILE Aires Chile CHILE the countries in the region. -

Sexual Selection and Extinction in Deer Saloume Bazyan
Sexual selection and extinction in deer Saloume Bazyan Degree project in biology, Master of science (2 years), 2013 Examensarbete i biologi 30 hp till masterexamen, 2013 Biology Education Centre and Ecology and Genetics, Uppsala University Supervisor: Jacob Höglund External opponent: Masahito Tsuboi Content Abstract..............................................................................................................................................II Introduction..........................................................................................................................................1 Sexual selection........................................................................................................................1 − Male-male competition...................................................................................................2 − Female choice.................................................................................................................2 − Sexual conflict.................................................................................................................3 Secondary sexual trait and mating system. .............................................................................3 Intensity of sexual selection......................................................................................................5 Goal and scope.....................................................................................................................................6 Methods................................................................................................................................................8 -

Online Resources Supplemental Material Supplementary Information
Hystrix (2019) — online resources Supplemental Material Supplementary Information Phylogeny and diversity of moose (Alces alces, Cervidae, Mammalia) revealed by complete mitochondrial genomes M. Świsłocka, M. Matosiuk, M. Ratkiewicz, A. Borkowska, M. Czajkowska, P. Mackiewicz Table S1: List of primer pairs used for PCR and sequencing of mitogenomes in Alces alces. Primer Primer sequence Position Tm [℃] Product [bp] Genes Primer source 12S-FW GGTAAATCTCGTGCCAGCCA 00295 57.3 712 <12s_rRNA> Fajardo et al., 2007† 12S-REV TCCAGTATGCTTACCTTGTTACGAC 01007 56.2 00871c_F TGCTTAGTTGAATTAGGCAATG 00872 51.3 1176 <12s_rRNA...tRNA-Val...16s_rRNA> Matosiuk et al., 2014‡ 02052c_R AGAGAACAAGTGATTATGCTACC 02048 52.2 01950c_F ACCTCCAGCATAACTAGTATTGG 01945 53.7 1455 <16s_rRNA...tRNA-Leu...ND1> Matosiuk et al., 2014‡ 03402c_R AATGGTCCTGCTGCATACTCTA 03400 55.2 03140c_F CTACGAGCAGTAGCTCAAACA 03138 54.1 1025 <ND1...tRNA-Ile...tRNA-Gln...tRNA-Met...ND2> Matosiuk et al., 2014‡ 04165c_R ACAGTTCATTGGCCTGAAAATA 04163 52.5 3910a_F CCTTCCCGTACTAATAAACC 03894 50.0 1519 <tRNA-Met...ND2...tRNA-Trp...tRNA-Ala...> This study 4300a_F2 TCATCAGGCCTAATTCTACT 04279 - <tRNA-Asn...tRNA-Cys...tRNA-Tyr...COX1> 5430a_R TATGCCTGCTCARGCACCAA 05413 56.0 COX1_F TCAGCCATTTTACCTATGTTCA 05315 51.7 826 <tRNA-Tyr...COX1> GenBank§ COX1_R ATRTAGCCAAARGGTTCTTTTT 06141 48.5 06060a_F TCTTTGGACACCCCGAAGTA 06039 55.2 991 <COX1...tRNA-Ser...tRNA-Asp...COX2> This study 07050a_R ATGGGGTAAGCCATATGAGG 07030 53.8 06090a_F TCGTAACATACTACTCAGGG 06099 50.2 1503 <COX1...tRNA-Ser...tRNA-Asp...COX2> This study -

A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname
Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname Editors: Leeanne E. Alonso and Trond H. Larsen 67 CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed RAP (Grensgebergte and Kasikasima) of Southeastern Suriname Bulletin of Biological Assessment 67 Editors: Leeanne E. Alonso and Trond H. Larsen CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION The RAP Bulletin of Biological Assessment is published by: Conservation International 2011 Crystal Drive, Suite 500 Arlington, VA USA 22202 Tel : +1 703-341-2400 www.conservation.org Cover photos: The RAP team surveyed the Grensgebergte Mountains and Upper Palumeu Watershed, as well as the Middle Palumeu River and Kasikasima Mountains visible here. Freshwater resources originating here are vital for all of Suriname. (T. Larsen) Glass frogs (Hyalinobatrachium cf. taylori) lay their -
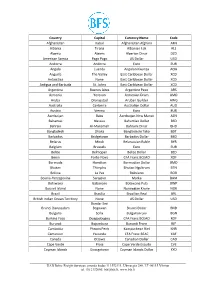
International Currency Codes
Country Capital Currency Name Code Afghanistan Kabul Afghanistan Afghani AFN Albania Tirana Albanian Lek ALL Algeria Algiers Algerian Dinar DZD American Samoa Pago Pago US Dollar USD Andorra Andorra Euro EUR Angola Luanda Angolan Kwanza AOA Anguilla The Valley East Caribbean Dollar XCD Antarctica None East Caribbean Dollar XCD Antigua and Barbuda St. Johns East Caribbean Dollar XCD Argentina Buenos Aires Argentine Peso ARS Armenia Yerevan Armenian Dram AMD Aruba Oranjestad Aruban Guilder AWG Australia Canberra Australian Dollar AUD Austria Vienna Euro EUR Azerbaijan Baku Azerbaijan New Manat AZN Bahamas Nassau Bahamian Dollar BSD Bahrain Al-Manamah Bahraini Dinar BHD Bangladesh Dhaka Bangladeshi Taka BDT Barbados Bridgetown Barbados Dollar BBD Belarus Minsk Belarussian Ruble BYR Belgium Brussels Euro EUR Belize Belmopan Belize Dollar BZD Benin Porto-Novo CFA Franc BCEAO XOF Bermuda Hamilton Bermudian Dollar BMD Bhutan Thimphu Bhutan Ngultrum BTN Bolivia La Paz Boliviano BOB Bosnia-Herzegovina Sarajevo Marka BAM Botswana Gaborone Botswana Pula BWP Bouvet Island None Norwegian Krone NOK Brazil Brasilia Brazilian Real BRL British Indian Ocean Territory None US Dollar USD Bandar Seri Brunei Darussalam Begawan Brunei Dollar BND Bulgaria Sofia Bulgarian Lev BGN Burkina Faso Ouagadougou CFA Franc BCEAO XOF Burundi Bujumbura Burundi Franc BIF Cambodia Phnom Penh Kampuchean Riel KHR Cameroon Yaounde CFA Franc BEAC XAF Canada Ottawa Canadian Dollar CAD Cape Verde Praia Cape Verde Escudo CVE Cayman Islands Georgetown Cayman Islands Dollar KYD _____________________________________________________________________________________________ -

Canada, Decembre 2008 Library and Bibliotheque Et 1*1 Archives Canada Archives Canada Published Heritage Direction Du Branch Patrimoine De I'edition
ORGANISATION SOCIALE, DYNAMIQUE DE POPULATION, ET CONSERVATION DU CERF HUEMUL (HIPPOCAMELUS BISULCUS) DANS LA PATAGONIE DU CHILI par Paulo Corti these presente au Departement de biologie en vue de l'obtention du grade de docteur es sciences (Ph.D.) FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE Sherbrooke, Quebec, Canada, decembre 2008 Library and Bibliotheque et 1*1 Archives Canada Archives Canada Published Heritage Direction du Branch Patrimoine de I'edition 395 Wellington Street 395, rue Wellington Ottawa ON K1A0N4 Ottawa ON K1A0N4 Canada Canada Your file Votre reference ISBN: 978-0-494-48538-5 Our file Notre reference ISBN: 978-0-494-48538-5 NOTICE: AVIS: The author has granted a non L'auteur a accorde une licence non exclusive exclusive license allowing Library permettant a la Bibliotheque et Archives and Archives Canada to reproduce, Canada de reproduire, publier, archiver, publish, archive, preserve, conserve, sauvegarder, conserver, transmettre au public communicate to the public by par telecommunication ou par Plntemet, prefer, telecommunication or on the Internet, distribuer et vendre des theses partout dans loan, distribute and sell theses le monde, a des fins commerciales ou autres, worldwide, for commercial or non sur support microforme, papier, electronique commercial purposes, in microform, et/ou autres formats. paper, electronic and/or any other formats. The author retains copyright L'auteur conserve la propriete du droit d'auteur ownership and moral rights in et des droits moraux qui protege cette these. this thesis. Neither the thesis Ni la these ni des extraits substantiels de nor substantial extracts from it celle-ci ne doivent etre imprimes ou autrement may be printed or otherwise reproduits sans son autorisation. -

Instituto Nacional De Pesquisas Da Amazônia – Inpa Programa De Pós-Graduação Em Ecologia
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA USO DE HABITAT E OCORRÊNCIA DE ROEDORES CAVIOMORFOS NA AMAZÔNIA CENTRAL, BRASIL FERNANDA DE ALMEIDA MEIRELLES Manaus, Amazonas Novembro, 2012 1 FERNANDA DE ALMEIDA MEIRELLES USO DE HABITAT E OCORRÊNCIA DE ROEDORES CAVIOMORFOS NA AMAZÔNIA CENTRAL, BRASIL ORIENTADOR: Dr. EDUARDO MARTINS VENTICINQUE Co-orientador: Dr. Torbjørn Haugaasen Dissertação apresentada ao Instituto Nacional de Pesquisas da Amazônia como parte dos requisitos para obtenção do título de Mestre em Biologia (Ecologia). Manaus, Amazonas Novembro, 2012 ii Banca examinadora não-presencial: Dra. Camila Righetto Cassano – Universidade Estadual de Santa Cruz Parecer: Aprovada com correções Dra. Maria Luisa da Silva Pinto Jorge – Universidade Estadual Paulista Júlio Mesquita Filho Parecer: Aprovada com correções Dr. José Manuel Vieria Fragoso – Universidade de Standfor, Califórnia (EUA) Parecer: Aprovada Banca examinadora presencial em 26 de novembro de 2012: Dr. Marcelo Gordo – Universidade Federal do Amazonas Parecer: Aprovada Dr. Paulo Estefano Dineli Bobrowiec – Instituto Nacional de Pesquisas da Amazônia Parecer: Aprovada Dr. Pedro Ivo Simões – Instituto Nacional de Pesquisas da Amazônia Parecer: Aprovada iii Sinopse: Estudei o uso de habitat e o efeito da distribuição de castanhais sobre a ocorrência de pacas, cotias e cutiaras na Reserva de Desenvolvimento Sustentável Piagaçu - Purus. Para isso distribuímos 107 câmeras-trap em locais com e sem exploração de castanha- do-Brasil. As variáveis ambientais, utilizadas como preditoras da ocorrência das espécies, foram medidas em campo ou extraídas de bases geográficas digitais. Os resultados indicam que as três espécies de roedores ocorrem preferencialmente em áreas com alta densidade de drenagem e distantes de comunidades humanas. -

Asymmetry and Karyotypic Relationships of Cervidae (Artiodactyla) Taxa
Punjab University Journal of Zoology 36(1): 71-79 (2021) https://dx.doi.org/10.17582/journal.pujz/2021.36.1.71.79 Research Article Karyotype Symmetry/ Asymmetry and Karyotypic Relationships of Cervidae (Artiodactyla) Taxa Halil Erhan Eroğlu Department of Biology, Faculty of Science and Art, Yozgat Bozok University, Yozgat, Turkey. Article History Received: February 26, 2019 Abstract | Karyotype asymmetry is one of the most widely used approaches in cytotaxonomic Revised: May 03, 2021 studies. The symmetry/asymmetry index (S/AI) is used to determine karyotype asymmetry in Accepted: May 18, 2021 higher animals and humans. The formula designed by the number of chromosome types is Published: June 13, 2021 S/AI = (1 × M) + (2 × SM) + (3 × A) + (4 × T) / 2n. The S/AI value varies from 1.0000 (full symmetric) to 4.0000 (full asymmetric). After a detailed literature review, the chromosomal Keywords data of 36 female species and 32 male species of family Cervidae were detected, namely (i) Artiodactyla, Cervidae, Kary- karyotype formulae, (ii) symmetry/asymmetry index values (iii) karyotype types. According otype, Phylogeny, Symmetry/ asymmetry index to the chromosomal data, two phylogenetic trees were formed. The phylogenetic trees were showed karyotypic relationships among the taxa. Novelty Statement | This is the first report on karyotype asymmetry of Cervidae. Karyotypic relationships are useful to infer processes of evolution and speciation. Karyotype asymmetry is an important parameter that helps to establish phylogenetic relationships among various species. To cite this article: Eroğlu, H.E., 2021. Karyotype symmetry/ asymmetry and karyotypic relationships of cervidae (Artiodactyla) taxa. Punjab Univ. J. Zool., 36(1): 71-79.