Hair Follicle Dermal Condensation Forms Via Fgf20 Primed Cell Cycle
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ARTICLES Fibroblast Growth Factors 1, 2, 17, and 19 Are The
0031-3998/07/6103-0267 PEDIATRIC RESEARCH Vol. 61, No. 3, 2007 Copyright © 2007 International Pediatric Research Foundation, Inc. Printed in U.S.A. ARTICLES Fibroblast Growth Factors 1, 2, 17, and 19 Are the Predominant FGF Ligands Expressed in Human Fetal Growth Plate Cartilage PAVEL KREJCI, DEBORAH KRAKOW, PERTCHOUI B. MEKIKIAN, AND WILLIAM R. WILCOX Medical Genetics Institute [P.K., D.K., P.B.M., W.R.W.], Cedars-Sinai Medical Center, Los Angeles, California 90048; Department of Obstetrics and Gynecology [D.K.] and Department of Pediatrics [W.R.W.], UCLA School of Medicine, Los Angeles, California 90095 ABSTRACT: Fibroblast growth factors (FGF) regulate bone growth, (G380R) or TD (K650E) mutations (4–6). When expressed at but their expression in human cartilage is unclear. Here, we deter- physiologic levels, FGFR3-G380R required, like its wild-type mined the expression of entire FGF family in human fetal growth counterpart, ligand for activation (7). Similarly, in vitro cul- plate cartilage. Using reverse transcriptase PCR, the transcripts for tivated human TD chondrocytes as well as chondrocytes FGF1, 2, 5, 8–14, 16–19, and 21 were found. However, only FGF1, isolated from Fgfr3-K644M mice had an identical time course 2, 17, and 19 were detectable at the protein level. By immunohisto- of Fgfr3 activation compared with wild-type chondrocytes and chemistry, FGF17 and 19 were uniformly expressed within the showed no receptor activation in the absence of ligand (8,9). growth plate. In contrast, FGF1 was found only in proliferating and hypertrophic chondrocytes whereas FGF2 localized predominantly to Despite the importance of the FGF ligand for activation of the resting and proliferating cartilage. -
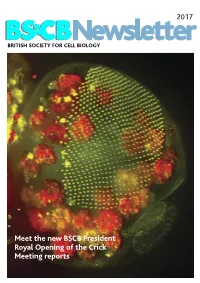
BSCB Newsletter 2017D
2017 BSCB Newsletter BRITISH SOCIETY FOR CELL BIOLOGY Meet the new BSCB President Royal Opening of the Crick Meeting reports 2017 CONTENTS BSCB Newsletter News 2 Book reviews 7 Features 8 Meeting Reports 24 Summer students 30 Society Business 33 Editorial Welcome to the 2017 BSCB newsletter. After several meeting hosted several well received events for our Front cover: years of excellent service, Kate Nobes has stepped PhD and Postdoc members, which we discuss on The head of a Drosophila pupa. The developing down and handed the reins over to me. I’ve enjoyed page 5. Our PhD and Postdoc reps are working hard compound eye (green) is putting together this years’ newsletter. It’s been great to make the event bigger and better for next year! The composed of several hundred simple units called ommatidia to hear what our members have been up to, and I social events were well attended including the now arranged in an extremely hope you will enjoy reading it. infamous annual “Pub Quiz” and disco after the regular array. The giant conference dinner. Members will be relieved to know polyploidy cells of the fat body (red), the fly equivalent of the The 2016 BSCB/DB spring meeting, organised by our we aren’t including any photos from that here. mammalian liver and adipose committee members Buzz Baum (UCL), Silke tissue, occupy a big area of the Robatzek and Steve Royle, had a particular focus on In this issue, we highlight the great work the BSCB head. Cells and Tissue Architecture, Growth & Cell Division, has been doing to engage young scientists. -

Ectodysplasin Target Gene Fgf20 Regulates Mammary Bud Growth and Ductal Invasion and Branching During Puberty
View metadata, citation and similar papersbroughtCORE at to core.ac.uk you by provided by Institute of Cancer Research Repository Ectodysplasin target gene Fgf20 regulates mammary bud growth and ductal invasion and branching during puberty Teresa Elo1,#, Päivi H. Lindfors1,#, Qiang Lan1 , Maria Voutilainen1, Ewelina Trela1, Claes Ohlsson2, Sung-Ho Huh3,^, David M. Ornitz3, Matti Poutanen4, Beatrice A. Howard5, Marja L. Mikkola1,* 1 Developmental Biology Program, Institute of Biotechnology, University of Helsinki, Finland 2 Center for Bone and Arthritis Research, Department of Internal Medicine, Institute of Medicine, Sahlgrenska Academy, University of Gothenburg, Sweden 3 Department of Developmental Biology, Washington University School of Medicine, St. Louis, Missouri, USA 4 Department of Physiology and Turku Center for Disease Modeling, Institute of Biomedicine, University of Turku, Turku, Finland 5 The Breast Cancer Now Toby Robins Research Centre, Division of Breast Cancer, the Institute of Cancer Research, London, United Kingdom #) these authors contributed equally ^) current address: Holland Regenerative Medicine Program and Department of Developmental Neuroscience, Munroe-Meyer Institute, University of Nebraska Medical Center, Omaha, Nebraska, USA *) Author for correspondence: Marja L. Mikkola Developmental Biology Program Institute of Biotechnology University of Helsinki P.O.B. 56 00014 Helsinki Finland e-mail: [email protected] phone: +358-2-941 59344 fax: +358-2-941 59366 1 Abstract Mammary gland development begins with the appearance of epithelial placodes that invaginate, sprout, and branch to form small arborized trees by birth. The second phase of ductal growth and branching is driven by the highly invasive structures called terminal end buds (TEBs) that form at ductal tips at the onset of puberty. -

FGF Signaling Network in the Gastrointestinal Tract (Review)
163-168 1/6/06 16:12 Page 163 INTERNATIONAL JOURNAL OF ONCOLOGY 29: 163-168, 2006 163 FGF signaling network in the gastrointestinal tract (Review) MASUKO KATOH1 and MASARU KATOH2 1M&M Medical BioInformatics, Hongo 113-0033; 2Genetics and Cell Biology Section, National Cancer Center Research Institute, Tokyo 104-0045, Japan Received March 29, 2006; Accepted May 2, 2006 Abstract. Fibroblast growth factor (FGF) signals are trans- Contents duced through FGF receptors (FGFRs) and FRS2/FRS3- SHP2 (PTPN11)-GRB2 docking protein complex to SOS- 1. Introduction RAS-RAF-MAPKK-MAPK signaling cascade and GAB1/ 2. FGF family GAB2-PI3K-PDK-AKT/aPKC signaling cascade. The RAS~ 3. Regulation of FGF signaling by WNT MAPK signaling cascade is implicated in cell growth and 4. FGF signaling network in the stomach differentiation, the PI3K~AKT signaling cascade in cell 5. FGF signaling network in the colon survival and cell fate determination, and the PI3K~aPKC 6. Clinical application of FGF signaling cascade in cell polarity control. FGF18, FGF20 and 7. Clinical application of FGF signaling inhibitors SPRY4 are potent targets of the canonical WNT signaling 8. Perspectives pathway in the gastrointestinal tract. SPRY4 is the FGF signaling inhibitor functioning as negative feedback apparatus for the WNT/FGF-dependent epithelial proliferation. 1. Introduction Recombinant FGF7 and FGF20 proteins are applicable for treatment of chemotherapy/radiation-induced mucosal injury, Fibroblast growth factor (FGF) family proteins play key roles while recombinant FGF2 protein and FGF4 expression vector in growth and survival of stem cells during embryogenesis, are applicable for therapeutic angiogenesis. Helicobacter tissues regeneration, and carcinogenesis (1-4). -

Reflections on the Historiography of Molecular Biology
Reflections on the Historiography of Molecular Biology HORACE FREELAND JUDSON SURELY the time has come to stop applying the word revolution to the rise of new scientific research programmes. Our century has seen many upheavals in scientific ideas--so many and so varied that the notion of scientific revolution has been stretched out of shape and can no longer be made to cover the processes of change characteristic of most sciences these past hundred years. By general consent, two great research pro- grammes arising in this century stand om from the others. The first, of course, was the one in physics that began at the turn of the century with quantum theory and relativity and ran through the working out, by about 1930, of quantum mechanics in its relativistic form. The trans- formation in physics appears to be thoroughly documented. Memoirs and biographies of the physicists have been written. Interviewswith survivors have been recorded and transcribed. The history has been told at every level of detail and difficulty. The second great programme is the one in biology that had its origins in the mid-1930s and that by 1970 had reached, if not a conclusion, a kind of cadence--a pause to regroup. This is the transformation that created molecular biology and latter-day biochemistry. The writing of its history has only recently started and is beset with problems. Accounting for the rise of molecular biology began with brief, partial, fugitive essays by participants. Biographies have been written of two, of the less understood figures in the science, who died even as the field was ripening, Oswald Avery and Rosalind Franklin; other scientists have wri:tten their memoirs. -

FGF20 and DKK1 Are Transcriptional Targets of Catenin and FGF20 Is
The EMBO Journal (2005) 24, 73–84 | & 2005 European Molecular Biology Organization | All Rights Reserved 0261-4189/05 www.embojournal.org THE EMBO JOJOURNALUR NAL FGF-20 and DKK1 are transcriptional targets of b-catenin and FGF-20 is implicated in cancer and development Mario N Chamorro1,4, Introduction Donald R Schwartz2,5, Alin Vonica3,5, Wnt proteins are secreted glycoproteins that bind and acti- Ali H Brivanlou3, Kathleen R Cho2 1, vate two classes of co-receptors, LDL-related proteins (LRPs) and Harold E Varmus * and members of the Frizzled protein family. Signaling in- 1Cancer Biology and Genetics Program, Sloan-Kettering Institute, itiated by Wnts and their receptors controls a wide variety of Varmus Laboratory, Memorial Sloan-Kettering Cancer Center, New York, cell processes, including cell fate specification, differentia- NY, USA, 2Department of Pathology, The University of Michigan Medical 3 tion, migration, and polarity (reviewed in Peifer and Polakis, School, Ann Arbor, MI, USA, The Laboratory of Vertebrate Embryology, b The Rockefeller University, New York, NY, USA and 4Cell Biology 2000). -Catenin is the major effector of the canonical Wnt Program, Cornell University, Weill Graduate School of Medical Sciences, signaling pathway. In the absence of Wnt, cytosolic b-catenin New York, NY, USA forms a complex with Axin and adenomatous polyposis coli (APC) proteins, and is rapidly degraded by the ubiquitina- b-catenin is the major effector of the canonical Wnt signal- tion–proteosome system. Wnt signaling inactivates the ing pathway. Mutations in components of the pathway that b-catenin destruction complex, so that b-catenin is stabilized, stabilize b-catenin result in augmented gene transcription accumulates in the cytoplasm and nucleus, and forms hetero- and play a major role in many human cancers. -

Front Matter (PDF)
<!< VOLUME 38 •NO. 4 CNREA8 •PP 877-1 179 April 1978 Nowyoucanhaveit bothways!i Thesensitivityyouneed todetectminorcomponents (o.oo50Dfullscale) PharmaciaDualPathMonitorUV-2 has a unique flow cell with two optical path-lengths for new versatility in UV-monitoring. Monitor absorbance quantitatively up to The Dual Path Monitor UV-2 has all the 20 OD units full scale with a sensitivity other features you expect of a high per of 0.005 OD units full scale at the same formance monitor: stability, cold room time, in the same run. operation convenience and compact de sign. For less-demanding applications you Monitor at 254 nm and/or 280 nm with can choose the Pharmacia Single Path two completely independent measuring Monitor UV-1 with a choice of 3 mm or systems. 10 mm flow cell and operation at 254 nm or 280 nm. Find out more about the practical advantages of column monitoring with the UV-2 and UV-1 Monitors. Ask about the Pharmacia Recorders too. Pharmacia Fine Chemicals Division of Pharmacia, Inc. Pharmacia Piscataway, New Jersey 08854 Phone (201)469-1222 Fine Chemicals COVER LEGEND. trate his attention on the study of living cells in vitro. His collaboration with Honor Fell resulted in the first key articles that initiated the concept and methods of organ culture (3). Honor B. Fell was born in 1900 and received her doctorate from the University of Edinburgh in 1924. She joined Strangeways the same year. Upon Strangeways' death the Research Hospital almost foundered, but after a difficult period it was rees tablished as the Strangeways Research Laboratory, with Dr. -
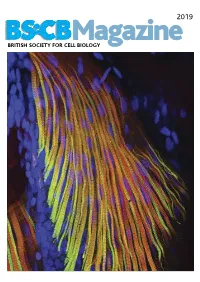
BSCB Newsletter 2019A:BSCB Aut2k7
2019 BSCB Magazine BRITISH SOCIETY FOR CELL BIOLOGY 2019 CONTENTS BSCB Magazine News 2 Book reviews 8 Features 9 Meeting Reports 21 Summer students 25 Society Business 32 Editorial Front cover: microscopic Welcome to the 2019 BSCB Magazine! This year Mustafa Aydogan (University of Oxford), as well as to structure of pectoral fin and Susana and Stephen are filling in for our Newsletter BSCB postdoc poster of the year winners Dr Anna hypaxial muscles of a zebrafish Editor Ann Wheeler. We hope you will enjoy this Caballe (University of Oxford) and Dr Agata Gluszek- Danio rerio larvae at four days year’s magazine! Kustusz (University of Edinburgh). post fertilization. The immunostaining highlights the This year we had a number of fantastic one day In 2019, we will have our jointly BSCB-BSDB organization of fast (red) and meetings sponsored by BSCB. These focus meetings Spring meeting at Warwick University from 7th–10th slow (green) myosins. All nuclei are great way to meet and discuss your science with April, organised by BSCB members Susana Godinho are highlighted in blue (hoechst). experts in your field and to strengthen your network of and Vicky Sanz-Moreno. The programme for this collaborators within the UK. You can read more about meeting, which usually provides a broad spectrum of these meetings in the magazine. If you have an idea themes, has a focus on cancer biology: cell for a focus one day meeting, check how to apply for migration/invasion, organelle biogenesis, trafficking, funding on page 4. Our ambassadors have also been cell-cell communication. -

Ectodysplasin Target Gene Fgf20 Regulates Mammary Bud Growth and Ductal Invasion and Branching During Puberty Teresa Elo University of Helsinki
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2017 Ectodysplasin target gene Fgf20 regulates mammary bud growth and ductal invasion and branching during puberty Teresa Elo University of Helsinki Päivi H. Lindfors University of Helsinki Qiang Lan University of Helsinki Maria Voutilainen University of Helsinki Ewelina Trela University of Helsinki See next page for additional authors Follow this and additional works at: https://digitalcommons.wustl.edu/open_access_pubs Recommended Citation Elo, Teresa; Lindfors, Päivi H.; Lan, Qiang; Voutilainen, Maria; Trela, Ewelina; Ohlsson, Claes; Huh, Sung-Ho; Ornitz, David M.; Poutanen, Matti; Howard, Beatrice A.; and Mikkola, Marja L., ,"Ectodysplasin target gene Fgf20 regulates mammary bud growth and ductal invasion and branching during puberty." Scientific Reports.7,. (2017). https://digitalcommons.wustl.edu/open_access_pubs/6000 This Open Access Publication is brought to you for free and open access by Digital Commons@Becker. It has been accepted for inclusion in Open Access Publications by an authorized administrator of Digital Commons@Becker. For more information, please contact [email protected]. Authors Teresa Elo, Päivi H. Lindfors, Qiang Lan, Maria Voutilainen, Ewelina Trela, Claes Ohlsson, Sung-Ho Huh, David M. Ornitz, Matti Poutanen, Beatrice A. Howard, and Marja L. Mikkola This open access publication is available at Digital Commons@Becker: https://digitalcommons.wustl.edu/open_access_pubs/6000 www.nature.com/scientificreports OPEN Ectodysplasin target gene Fgf20 regulates mammary bud growth and ductal invasion and branching Received: 28 December 2016 Accepted: 18 May 2017 during puberty Published: xx xx xxxx Teresa Elo1, Päivi H. Lindfors1, Qiang Lan1, Maria Voutilainen1, Ewelina Trela1, Claes Ohlsson2, Sung-Ho Huh3,6, David M. -

CHEMICAL HERITAGE FOUNDATION GERALD WEISSMANN the Pew
CHEMICAL HERITAGE FOUNDATION GERALD WEISSMANN The Pew Scholars Program in the Biomedical Sciences Transcript of an Interview Conducted by Arthur Daemmrich at The Marine Biological Laboratory Woods Hole, Massachusetts on 2 August 2007 (With Subsequent Corrections and Additions) Gerald Weissmann ACKNOWLEDGEMENT This oral history is part of a series supported by a grant from the Pew Charitable Trusts based on the Pew Scholars Program in the Biomedical Sciences. This collection is an important resource for the history of biomedicine, recording the life and careers of young, distinguished biomedical scientists and of Pew Biomedical Scholar Advisory Committee members. This oral history is made possible through the generosity of This interview has been designated as Free Access. One may view, quote from, cite, or reproduce the oral history with the permission of CHF. Please note: Users citing this interview for purposes of publication are obliged under the terms of the Chemical Heritage Foundation Oral History Program to credit CHF using the format below: Gerald Weissmann, interview by Arthur Daemmrich at The Marine Biological Laboratory, Woods Hole, Massachusetts, 2 August 2007 (Philadelphia: Chemical Heritage Foundation, Oral History Transcript # 0371). Chemical Heritage Foundation Oral History Program 315 Chestnut Street Philadelphia, Pennsylvania 19106 The Chemical Heritage Foundation (CHF) serves the community of the chemical and molecular sciences, and the wider public, by treasuring the past, educating the present, and inspiring the future. CHF maintains a world-class collection of materials that document the history and heritage of the chemical and molecular sciences, technologies, and industries; encourages research in CHF collections; and carries out a program of outreach and interpretation in order to advance an understanding of the role of the chemical and molecular sciences, technologies, and industries in shaping society. -

Expression of Fgfs During Early Mouse Tongue Development
Gene Expression Patterns 20 (2016) 81e87 Contents lists available at ScienceDirect Gene Expression Patterns journal homepage: http://www.elsevier.com/locate/gep Expression of FGFs during early mouse tongue development * Wen Du a, b, Jan Prochazka b, c, Michaela Prochazkova b, c, Ophir D. Klein b, d, a State Key Laboratory of Oral Diseases, West China Hospital of Stomatology, Sichuan University, Chengdu, Sichuan, 610041, China b Department of Orofacial Sciences and Program in Craniofacial Biology, University of California San Francisco, San Francisco, CA 94143, USA c Laboratory of Transgenic Models of Diseases, Institute of Molecular Genetics of the ASCR, v.v.i., Prague, Czech Republic d Department of Pediatrics and Institute for Human Genetics, University of California San Francisco, San Francisco, CA 94143, USA article info abstract Article history: The fibroblast growth factors (FGFs) constitute one of the largest growth factor families, and several Received 29 September 2015 ligands and receptors in this family are known to play critical roles during tongue development. In order Received in revised form to provide a comprehensive foundation for research into the role of FGFs during the process of tongue 13 December 2015 formation, we measured the transcript levels by quantitative PCR and mapped the expression patterns by Accepted 29 December 2015 in situ hybridization of all 22 Fgfs during mouse tongue development between embryonic days (E) 11.5 Available online 31 December 2015 and E14.5. During this period, Fgf5, Fgf6, Fgf7, Fgf9, Fgf10, Fgf13, Fgf15, Fgf16 and Fgf18 could all be detected with various intensities in the mesenchyme, whereas Fgf1 and Fgf2 were expressed in both the Keywords: Tongue epithelium and the mesenchyme. -

Kif Liakath-Ali Postdoctoral Research Fellow, Molecular and Cellular Physiology
Kif Liakath-Ali Postdoctoral Research Fellow, Molecular and Cellular Physiology Bio BIO Dr Liakath-Ali holds a PhD degree in molecular genetics from the University of Cambridge, UK. He carried out his doctoral and a brief post-doctoral research under the supervision of Professor Fiona Watt at Cambridge and King’s College London. While in Watt lab, he conducted a first, large-scale tissue-specific phenotype screen on hundreds of knockout mice and discovered many novel genes that are essential for mammalian skin function. He further elucidated the mechanistic roles of sphingolipid and a ribosome-rescue pathway in epidermal stem cell function. He has published many papers in the area of skin biology and won several awards, including, most recently a long-term fellowship from the European Molecular Biology Organization (EMBO) and postdoctoral/principal investigator grant from the Larry L Hillblom Foundation. Dr Liakath-Ali obtained his bachelor and master degree in Zoology from Jamal Mohamed College (Bharathidasan University), Trichy, India. He further specialized in human genetics and obtained an MPhil from the University of Madras, India. He went on to work at various capacities at the Centre for DNA Fingerprinting and Diagnostics, Hyderabad, India, Institute of Human Genetics, University of Göttingen, Germany and the Wellcome Sanger Institute, Cambridge, UK. Dr Liakath-Ali also holds a degree equivalent (Associateship of King’s College (AKC) in Philosophy, Ethics and Theology, awarded by King's College London, UK. It is perhaps these combinations of diverse backgrounds and training that led Dr Liakath-Ali to develop an interest in fundamental questions in neuroscience.