JBLM Streaked Horned Lark Surveys and Monitoring 2016
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JBLM Lark Monitoring Final Report 2015 W911S8-14-2-0026 W911S8-15-2-0001 CNLM Task Orders #G1108, G1118 April 2016
Habitat and Species Cooperative Restoration Program Joint Base Lewis-McChord Center for Natural Lands Management JBLM Lark Monitoring Final Report 2015 W911S8-14-2-0026 W911S8-15-2-0001 CNLM Task Orders #G1108, G1118 April 2016 Submitted to: Joint Base Lewis-McChord Fish and Wildlife Program Submitted by: Adrian Wolf, Gary Slater and Hannah Anderson Center for Natural Lands Management 120 Union Avenue Southeast Olympia WA, 98501 Phone: 360-584-2538 Joint Base Lewis-McChord is a key military installation and the most important conservation area in the Puget Trough region. The Center for Natural Lands Management strives to assist Joint Base Lewis-McChord in the conservation of its natural resources within the framework of the military training mandate. Joint Base Lewis- McChord and its conservation partners have shared interests because: Healthy natural ecosystems are essential for realistic and sustainable training lands. Rare species recovery throughout the region reduces the burden of recovery on any single landowner or site. Pest plants harm natural areas and reduce their suitability for military training. Page 2 Table of Contents Project Highlights ............................................................................................................................ 5 1.0 Introduction ............................................................................................................................. 6 1.1 Goals and Objectives ........................................................................................................... -

Specifications and Contract Documents
SPECIFICATIONS AND CONTRACT DOCUMENTS Killeen, Texas Robert Gray Army Airfield (RGAAF) Security Surveillance Improvements at Robert Gray Army Airfield City of Killeen RFP No. 19-10 ISSUED FOR RFP Garver Project No. 17181082 Project Sponsors: Prepared For: City of Killeen November 6, 2018 This page intentionally left blank Security Surveillance Improvements at RGAAF Project No. 17181082 Issued for RFP November 6, 2018 SECTION I. PROPOSAL REQUIREMENTS Security Surveillance Improvements at RGAAF Project No. 17181082 Issued for RFP November 6, 2018 This page intentionally left blank Security Surveillance Improvements at RGAAF Project No. 17181082 Issued for RFP November 6, 2018 SECTION 000001 CERTIFICATIONS SECURITY SURVEILLANCE IMPROVEMENTS AT ROBERT GRAY ARMY AIRFIELD PROJECT GARVER PROJECT NO. 17181082 CITY OF KILLEEN RFP NO. 19-10 I hereby certify that the applicable portions of this project plans and specifications were prepared by me or under my direct supervision and that I am a duly Licensed Engineer under the laws of the State of Texas. APPLICABLE DIVISION OR SEAL AND SIGNATURE PROJECT RESPONSIBILITY Derek Mayo, P.E. General Plans and Specifications Digitally Signed: Matthew LeMay, P.E. Electrical Plans and Specifications Digitally Signed: Security Surveillance Improvements at RGAAF Project No. 17181082 Issued for RFP 000001-1 November 6, 2018 SECTION 000001 CERTIFICATIONS APPLICABLE DIVISION OR SEAL AND SIGNATURE PROJECT RESPONSIBILITY Michael A. Guzik, P.E. CTI Telecom Plans and Specifications Digitally Signed GARVER, LLC CERTIFICATE OF AUTHORIZATION: TX ENGINEERING REGISTRATION NO. F-5713 TX ARCHITECTURAL REGISTRATION NO. 21507 CTI CERTIFICATE OF AUTHORIZATION: TX REGISTERED CONSULTING FIRM B-17368 Security Surveillance Improvements at RGAAF Project No. -

Unmanned Aerial Systems (UAS) Market Overview
Unmanned Aerial Systems (UAS) Market Overview July 18, 2012 Contents . Definitions . Facts & Figures . UAS Budgets and Spending . Trends and Challenges . Future of Unmanned Aerial Systems . Summary and Recommendations . Appendix A: DoD UAS Acquisition Costs . Appendix B: DoD Current and Future Domestic UAS Locations 2 7/18/2012 ©2012 Deltek, Inc. All Rights Reserved Definitions . Unmanned Aerial Systems (UAS) . Refers to systems whose components include the necessary equipment, network, and personnel to control an unmanned aircraft . UAS is a broader term that includes equipment, networks, and personnel in addition to Unmanned Aerial Vehicles. Unmanned Aerial Vehicle (UAV) . Refers to a powered aerial vehicle that does not carry a human operation, uses aerodynamic forces to provide vehicle life, can fly autonomously or be piloted remotely, can be expendable or recoverable, and can carry a lethal or nonlethal payload . Commonly known as “drones”. In practice, the terms UAS and UAV are often used interchangeably Source: CRS Report R41284, “Intelligence, Surveillance, and Reconnaissance (ISR) Acquisition: Issues for Congress,” December 27, 2011. 3 7/18/2012 ©2012 Deltek, Inc. All Rights Reserved Facts & Figures . Dept. of Defense (DoD) spending on UAS has increased from $284 million in FY00 to $3.9 billion in FY12 . DoD’s unmanned aircraft inventory increased more than 40-fold from 167 aircraft in 2002 to nearly 7,500 in 2010 . In 2009, DoD completed almost 500,000 UAS flight hours just in support of Operation Enduring Freedom and Operation Iraqi Freedom . In May 2010, unmanned systems surpassed one million flight hours . In November 2010 unmanned systems achieved one million combat hours Sources: CRS Report R41284, “Intelligence, Surveillance, and Reconnaissance (ISR) Acquisition: Issues for Congress,” December 27, 2011; CRS Report R42136, “Unmanned Aerial Systems,” January 3, 2012; Dept. -

Fort Hood Noise Study
130 TRANSPORTATION RESEARCH RECORD 1312 Fort Hood Noise Study RICHARD M. LETTY At the request' of the U.S. Army Corp of Engineer , Fort WortJ1 measuring airfield noise and impulsive weapon-firing blast District Office, an in tallation compatible-use zone (ICUZ) noise noise, it was decided that the noise monitoring would be study was prepared for Fort Hood, Texa . The purpose f this performed in two phases. The Phase I noise measurements study was to addres the n i e impact from military training ac focused on airfield noise. A total of nine noise measurement tivity conducted at Fort Hood. The major component of this Fort Hood I UZ noise study wa a comprehensive long-term noise locations were selected: two in the vicinity of RGAAF, four monitoring program and the use of computer modeling to develop around HAAF, and three along the various flight corridors noi e contour to iden tify noise-impacted areas. Noise measure leading to and from the Fort Hood Army Installation. The ments were obtained at a total of 17 noise measurement locations: Phase II noise measurements focused on the blast noise from 9 airfield noi e monitoring ·itc , and 8 weapon-fi ring blast noise artillery and weapon-firing activity on the various ranges at monitoring site . Because of the day-to-day variations in military Fort Hood. The purpose of the noise monitoring program was training activity, it was determined that 60 days of noise data at ach of the 17 noise-m nitoring sites would be useful in under not only to define the actual noise levels from airfield and standing long-term airfield and weapon-firing blast noise levels. -
Alaska Post Newspaper
FREE RECYCLED an edition of the Recycled material is used in the making of our ALASKA POST newsprint The Interior Military News Connection Vol. 8, No. 37 Fort Wainwright, Alaska September 15, 2017 Alaska, California rescue squadrons unite to save lives in Southeast Texas Staff Sgt. Balinda O’Neal Dresel 249th Airlift Squadron bound for Alaska National Guard Public Moffett Federal Airfield, Calif., Affairs to pick up two HH-60 Pave Hawk helicopters and aircrews from the Air National Guardsmen from 129th Rescue Squadron. They Alaska’s Chugach Mountains and the arrived in Fort Hood in the early- heart of California’s Silicon Valley morning hours of Aug. 29 and spent last week in the flooded cities of began moving equipment to a Southeast Texas, with one mission— staging area on Gray Army Airfield to save lives. where they were later paired Aircrews, combat rescue officers, with search and rescue personnel pararescuemen and support personnel from California’s 131st Rescue from the Alaska Air National Guard’s Squadron. 176th Wing and members of the “One of the greatest aspects California Air National Guard’s of working with other pararescue 129th Rescue Wing joined more than teams is that we can come together 18,000 National Guard personnel under a common thread and who responded to the call to assist train and work as a joint force,” with Hurricane Harvey humanitarian explained Senior Master Sgt. disaster relief operations. The Airmen Brandon Stuemke, a pararescueman left home Aug. 28 to help their with the 212th Rescue Squadron. neighbors in Texas still needing relief “It allowed for us to seamlessly Air National Guard search and rescue personnel from Alaska and California conduct water and evacuation. -

National Plan of Integrated Airport Systems (NPIAS) (2011-2015)
NPIAS 2011-2015 Illustrated by GRA, Incorporated Federal Aviation Administration U.S. Department of Transportation National Plan of Integrated Airport Systems (NPIAS) (2011-2015) Report of the Secretary of Transportation to the United States Congress Pursuant to Section 47103 of Title 49, United States Code The NPIAS 2011-2015 report is available online at http://www.faa.gov/airports/planning_capacity/npias/reports Table of Contents EXECUTIVE SUMMARY .............................................................................................................. VI Development Estimates .......................................................................................................... vii Estimates by Airport Type......................................................................................... viii Estimates by Type of Development...............................................................................x Status of the Industry .............................................................................................................. xii CHAPTER 1: SYSTEM COMPOSITION.......................................................................................1 Overview....................................................................................................................................1 U.S. Department of Transportation................................................................................2 Federal Aviation Administration ...................................................................................2 -

EXHIBIT 2 Recovery Outline for the Streaked Horned Lark
EXHIBIT 2 Recovery Outline for the Streaked Horned Lark U.S. Fish & Wildlife Service Recovery Outline for the Streaked Horned Lark (Eremophila alpestris strigata) Photo: D. Leonard, USFWS Common Name Streaked horned lark Scientific Name Eremophila alpestris strigata Listing Status and Date Threatened; October 3, 2013 (78 FR 61452) Critical Habitat and Date Designated; October 3, 2013 (78 FR 61506) Lead Agency/Region U.S. Fish and Wildlife Service, Region 1 Lead Field Office Oregon Fish and Wildlife Office 2600 SE 98th Avenue, Suite 100 Portland, Oregon 97266 (503) 231-6179 Lead Biologist Cat Brown (503) 231-6179, [email protected] Purpose of the Recovery Outline: This document lays out a preliminary course of action for the survival and recovery of the streaked horned lark. It is meant to serve as interim guidance to direct recovery efforts and inform consultation and permitting activities until a comprehensive draft recovery plan has been completed. Recovery outlines are intended primarily for internal use by the U.S. Fish and Wildlife Service (Service), and formal public participation will be invited upon the release of the draft recovery plan. However, we will consider any new information or comments that members of the public may wish to offer in response to this outline during the recovery planning process. For more information on Federal survival and recovery efforts for the streaked horned lark, or to provide additional comments, interested parties may contact 1 Recovery Outline for the Streaked Horned Lark the lead biologist for this species, Cat Brown, at the above address, telephone number, or e-mail. -
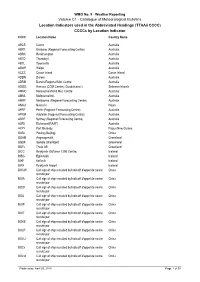
Catalogue of Meteorological Bulletins Location Indicators Used in the Abbreviated Headings (Ttaaii CCCC) Ccccs by Location Indicator CCCC Location Name Country Name
WMO No. 9 - Weather Reporting Volume C1 - Catalogue of Meteorological Bulletins Location Indicators used in the Abbreviated Headings (TTAAii CCCC) CCCCs by Location Indicator CCCC Location Name Country Name ABCS Cairns Australia ABRF Brisbane (Regional Forecasting Centre) Australia ABRK Rockhampton Australia ABTD Thursday I. Australia ABTL Townsville Australia ABWP Weipa Australia ACCC Cocos Island Cocos Island ADDN Darwin Australia ADRM Darwin/Regional Met. Centre Australia AGGG Honiara (COM Centre), Guadalcanal I. Solomon Islands AMMC Melbourne/World Met. Centre Australia AMML Melbourne/Intl. Australia AMRF Melbourne (Regional Forecasting Centre) Australia ANAU Nauru Is. Nauru APRF Perth (Regional Forecasting Centre) Australia APRM Adelaide (Regional Forecasting Centre) Australia ASRF Sydney (Regional Forecasting Centre) Australia ASRI Richmond(RAAF) Australia AYPY Port Moresby Papua New Guinea BABJ Peking (Beijing) China BGAM Angmagssalik Greenland BGSF Søndre Strømfjord Greenland BGTL Thule AB Greenland BICC Reykjavik (Gufunes COM Centre) Iceland BIEG Egilsstadir Iceland BIKF Keflavík Iceland BIRK Reykjavík Airport Iceland BOCW Call sign of ship recruited by/Indicatif d'appel de navire China recruté par BOIA Call sign of ship recruited by/Indicatif d'appel de navire China recruté par BOID Call sign of ship recruited by/Indicatif d'appel de navire China recruté par BOIJ Call sign of ship recruited by/Indicatif d'appel de navire China recruté par BOIP Call sign of ship recruited by/Indicatif d'appel de navire China recruté par BOIT -

Department of the Interior
Vol. 78 Thursday, No. 192 October 3, 2013 Part III Department of the Interior Fish and Wildlife Service 50 CFR Part 17 Endangered and Threatened Wildlife and Plants; Designation of Critical Habitat for Taylor’s Checkerspot Butterfly and Streaked Horned Lark; Final Rule VerDate Mar<15>2010 18:36 Oct 02, 2013 Jkt 232001 PO 00000 Frm 00001 Fmt 4717 Sfmt 4717 E:\FR\FM\03OCR3.SGM 03OCR3 tkelley on DSK3SPTVN1PROD with RULES3 61506 Federal Register / Vol. 78, No. 192 / Thursday, October 3, 2013 / Rules and Regulations DEPARTMENT OF THE INTERIOR by appointment, at the Washington Fish Washington; and in Benton County in and Wildlife Office (see FOR FURTHER Oregon. Fish and Wildlife Service INFORMATION CONTACT). Any additional • Approximately 4,629 ac (1,873 ha) tools or supporting information that we in two units for the streaked horned lark 50 CFR Part 17 developed for this critical habitat in Grays Harbor, Pierce, Pacific, and [Docket No. FWS–R1–ES2013–0009; designation will also be available at the Wahkiakum Counties in Washington; 4500030114] Fish and Wildlife Service Web site and and in Clatsop, Columbia, Marion, Polk, field office set out above, and may also and Benton Counties in Oregon. RIN 1081–AZ36 be included at http:// We have prepared an economic www.regulations.gov. analysis of the designation of critical Endangered and Threatened Wildlife habitat. We have prepared an analysis FOR FURTHER INFORMATION CONTACT: Ken and Plants; Designation of Critical of the probable economic impacts of the Berg, Manager, U.S. Fish and Wildlife Habitat for Taylor’s Checkerspot critical habitat designations and related Service, Washington Fish and Wildlife Butterfly and Streaked Horned Lark factors. -

Recovery Outline for the Streaked Horned Lark
Recovery Outline for the Streaked Horned Lark U.S. Fish & Wildlife Service Recovery Outline for the Streaked Horned Lark (Eremophila alpestris strigata) Photo: D. Leonard, USFWS Common Name Streaked horned lark Scientific Name Eremophila alpestris strigata Listing Status and Date Threatened; October 3, 2013 (78 FR 61452) Critical Habitat and Date Designated; October 3, 2013 (78 FR 61506) Lead Agency/Region U.S. Fish and Wildlife Service, Region 1 Lead Field Office Oregon Fish and Wildlife Office 2600 SE 98th Avenue, Suite 100 Portland, Oregon 97266 (503) 231-6179 Lead Biologist Cat Brown (503) 231-6179, [email protected] Purpose of the Recovery Outline: This document lays out a preliminary course of action for the survival and recovery of the streaked horned lark. It is meant to serve as interim guidance to direct recovery efforts and inform consultation and permitting activities until a comprehensive draft recovery plan has been completed. Recovery outlines are intended primarily for internal use by the U.S. Fish and Wildlife Service (Service), and formal public participation will be invited upon the release of the draft recovery plan. However, we will consider any new information or comments that members of the public may wish to offer in response to this outline during the recovery planning process. For more information on Federal survival and recovery efforts for the streaked horned lark, or to provide additional comments, interested parties may contact 1 Recovery Outline for the Streaked Horned Lark the lead biologist for this species, Cat Brown, at the above address, telephone number, or e-mail. Scope of Recovery and Available Information: The scope of this effort is for a single species, the streaked horned lark. -

Preparing Busy General Aviation Airports for Next Generation Technologies
Preparing Busy General Aviation Airports for Next Generation Technologies May 2013 2013PSRC Preparing Busy Airports for NextGen Technologies The preparation of this report was financed in part through a planning grant from the Federal Aviation Administration (FAA). The grant was provided under Section 505 of the Airport and Airway Improvement Act of 1982, as amended by the Aviation Safety and Capacity Expansion Acts of 1987 and 1990, and the Airport Noise and Capacity Act of 1990. The contents do not necessarily reflect the official views or policy of the FAA. Acceptance of this report by the FAA under the terms of the scope of work and the grant agreement does not in any way constitute a commitment on the part of the United States to participate in any policy, plan, or development depicted herein, nor does it indicate that the proposed policy, plan, or development is environmentally acceptable in accordance with appropriate public laws. For additional information or to obtain copies of the Preparing Busy General Aviation Airports for NextGen Technologies report, go to www.psrc.org/transportation/airtrans/nextgen/ or contact the Puget Sound Regional Council’s Information Center at 1-206-464-7532 or [email protected] PSRC Preparing Busy Airports for NextGen Technologies NextGen Project Advisory Committee (PAC) Members Agency Bill Ayer CEO, Alaska Air Group (retired) Sarah Dalton FAA - Airports Division Manager Carolyn Read FAA - Northwest Mountain Region Charlie Howard Puget Sound Regional Council Chris Pomeroy WSDOT Aviation (former) Chuck Kegley Advanced Aviation Services, Inc. - PNBAA Board Deb Wallace Pierce County Airport and Ferry Administrator Dick Taylor Boeing Test Pilot (retired) Fred Mitchell FAA - Western Flight Procedures Office Gary Molyneaux King County International Airport (Boeing Field) Gene McBrayer Museum of Flight, Exxon Corp. -

The Trans-Texas Corridor and the Texas Airport System: Opportunities and Challenges (FHWA/TX-06/0-4644-1)
Technical Report Documentation Page 1. Report No. 2. Government 3. Recipient’s Catalog No. FHWA/TX-06/0-4644-1 Accession No. 4. Title and Subtitle 5. Report Date The Trans-Texas Corridor and the Texas Airport System: October 2004; Revised May 2006 Opportunities and Challenges 6. Performing Organization Code 7. Author(s) 8. Performing Organization Report No. Kelsey A. Thompson, Michael S. Bomba, C. Michael Walton, 0-4644-1 Jordan E. Botticello 9. Performing Organization Name and Address 10. Work Unit No. (TRAIS) Center for Transportation Research 11. Contract or Grant No. The University of Texas at Austin 0-4644 3208 Red River, Suite 200 Austin, TX 78705-2650 12. Sponsoring Agency Name and Address 13. Type of Report and Period Covered Texas Department of Transportation Technical Report Research and Technology Implementation Office August 2003–August 2004 P.O. Box 5080 Austin, TX 78763-5080 14. Sponsoring Agency Code 15. Supplementary Notes Project performed in cooperation with the Texas Department of Transportation and the Federal Highway Administration. Project Title: Evaluation and Integration of Texas Airports into the Trans-Texas Corridor 16. Abstract The proposed Trans-Texas Corridor (TTC) will allow for faster and safer movement of people and goods throughout Texas, relieve congestion on existing roadways, divert hazardous materials away from urban areas, and stimulate economic growth and development along its path. However, to become fully integrated with the Texas transportation network, the TTC must also consider connections with the state’s extensive airport system. While the TTC could produce significant opportunities for commercial services and general aviation airports, many of its planners and engineers are not familiar with the special land-use and connectivity needs of airports.