Animal Traditions: Behavioural Inheritance in Evolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Introduction to Genetics Bios 225
INTRODUCTION TO GENETICS BIOS 225 Course Description This course introduces the student to the basic concepts of inheritance, populations, mutations, and techniques used to assess each of these. Credit: 2 credits Repeatable: No Course Structure The course will be presented in different formats: Lectures with PowerPoints, self-directed learning, discussions and student assignments etc. Competencies This course emphasizes competencies to enhance skills essential for a future health care professional. • Knowledge o Demonstrate content knowledge and skills in foundational courses required by biomedical professionals o Demonstrate information literacy o Demonstrate quantitative reasoning o Demonstrate longitudinal learning through coursework • Critical Thinking o Develop the skills of self-reflection and peer assessment to improve personal performance. o Demonstrate the ability to analyze literature and written material o Demonstrate the ability to distinguish between well-reasoned and poorly reasoned arguments • Communication Skills o Demonstrate effective presentation skills to faculty and peers. o Demonstrate effective listening skills o Demonstrate effective written communication 1 Objectives: Upon completion of BIOS 225 course, the student should be able to describe: • The structure and function of purines, pyrimidines, nucleosides and nucleotides • The structure and functions of nucleic acids (DNA and RNA) • The chromosome anatomy and human karyotypes • The concepts of prokaryotic and eukaryotic DNA replication • The concepts of prokaryotic and eukaryotic RNA transcription and post-transcriptional modifications • The concepts of prokaryotic and eukaryotic protein translation and post-translational modifications • The regulations of prokaryotic and eukaryotic gene expressions • The process of genomic, chromosomal and gene mutations; and its repair mechanism • The Mendel’ hypothesis and molecular mechanisms of genetic inheritance Schedule: Dates and times to be posted at the beginning of the term on the online calendar. -

THE NAKED APE By
THE NAKED APE by Desmond Morris A Bantam Book / published by arrangement with Jonathan Cape Ltd. PRINTING HISTORY Jonathan Cape edition published October 1967 Serialized in THE SUNDAY MIRROR October 1967 Literary Guild edition published April 1969 Transrvorld Publishers edition published May 1969 Bantam edition published January 1969 2nd printing ...... January 1969 3rd printing ...... January 1969 4th printing ...... February 1969 5th printing ...... June1969 6th printing ...... August 1969 7th printing ...... October 1969 8th printing ...... October 1970 All rights reserved. Copyright (C 1967 by Desmond Morris. This book may not be reproduced in whole or in part, by mitneograph or any other means, without permission. For information address: Jonathan Cape Ltd., 30 Bedford Square, London Idi.C.1, England. Bantam Books are published in Canada by Bantam Books of Canada Ltd., registered user of the trademarks con silting of the word Bantam and the portrayal of a bantam. PRINTED IN CANADA Bantam Books of Canada Ltd. 888 DuPont Street, Toronto .9, Ontario CONTENTS INTRODUCTION, 9 ORIGINS, 13 SEX, 45 REARING, 91 EXPLORATION, 113 FIGHTING, 128 FEEDING, 164 COMFORT, 174 ANIMALS, 189 APPENDIX: LITERATURE, 212 BIBLIOGRAPHY, 215 ACKNOWLEDGMENTS This book is intended for a general audience and authorities have therefore not been quoted in the text. To do so would have broken the flow of words and is a practice suitable only for a more technical work. But many brilliantly original papers and books have been referred to during the assembly of this volume and it would be wrong to present it without acknowledging their valuable assistance. At the end of the book I have included a chapter-by-chapter appendix relating the topics discussed to the major authorities concerned. -

Fish Cognition: a Primate's Eye View
Anim Cogn (2002) 5:1–13 DOI 10.1007/s10071-001-0116-5 REVIEW Redouan Bshary · Wolfgang Wickler · Hans Fricke Fish cognition: a primate’s eye view Received: 8 February 2001 / Accepted after revision: 6 September 2001 / Published online: 20 November 2001 © Springer-Verlag 2001 Abstract We provide selected examples from the fish lit- erature of phenomena found in fish that are currently be- Introduction ing examined in discussions of cognitive abilities and evolution of neocortex size in primates. In the context of Recently increased interest in cognitive aspects of the be- social intelligence, we looked at living in individualised haviour of animals is connected with advances in the de- groups and corresponding social strategies, social learning velopment of both theoretical frameworks and methodol- and tradition, and co-operative hunting. Regarding envi- ogy. Cognitive psychologists have adopted a more evolu- ronmental intelligence, we searched for examples concern- tionary approach to cognitive research (see historical re- ing special foraging skills, tool use, cognitive maps, mem- view in Kamil 1998). The major contribution of primatol- ory, anti-predator behaviour, and the manipulation of the ogy to cognitive psychology was arguably the develop- environment. Most phenomena of interest for primatolo- ment of the social intelligence hypothesis (Humphrey 1976; gists are found in fish as well. We therefore conclude that Byrne and Whiten 1988; Dunbar 1992), which states that more detailed studies on decision rules and mechanisms the evolution of cognitive skills together with a large neo- are necessary to test for differences between the cognitive cortex in primates was caused by the social complexity abilities of primates and other taxa. -

Taking Turns: Bridging the Gap Between Human and Animal Communication
This is a repository copy of Taking turns: bridging the gap between human and animal communication. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/150857/ Version: Published Version Article: Pika, Simone, Wilkinson, Ray, Kendrick, Kobin H. orcid.org/0000-0002-6656-1439 et al. (1 more author) (2018) Taking turns: bridging the gap between human and animal communication. Proceedings of the royal society b-Biological sciences. 20180598. ISSN 1471-2954 https://doi.org/10.1098/rspb.2018.0598 Reuse This article is distributed under the terms of the Creative Commons Attribution (CC BY) licence. This licence allows you to distribute, remix, tweak, and build upon the work, even commercially, as long as you credit the authors for the original work. More information and the full terms of the licence here: https://creativecommons.org/licenses/ Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Downloaded from http://rspb.royalsocietypublishing.org/ on July 30, 2018 Taking turns: bridging the gap between rspb.royalsocietypublishing.org human and animal communication Simone Pika1,2, Ray Wilkinson3, Kobin H. Kendrick4 and Sonja C. Vernes5,6 1Department of Primatology, Max Planck Institute for Evolutionary Anthropology, Deutscher Platz 6, 04103 Leipzig, Germany Review 2Department of Comparative Biocognition, Institute of Cognitive Science, University of Osnabru¨ck, Osnabru¨ck, Germany Cite this article: Pika S, Wilkinson R, 3Department of Human Communication Sciences, University of Sheffield, Sheffield, UK Kendrick KH, Vernes SC. -

Law in Sociobiological Perspective
Florida State University Law Review Volume 5 Issue 2 Article 2 Spring 1977 Law in Sociobiological Perspective Dr. Margaret Gruter Stanford University, Program in Human Biology Follow this and additional works at: https://ir.law.fsu.edu/lr Part of the Law and Society Commons Recommended Citation Dr. Margaret Gruter, Law in Sociobiological Perspective, 5 Fla. St. U. L. Rev. 181 (1977) . https://ir.law.fsu.edu/lr/vol5/iss2/2 This Article is brought to you for free and open access by Scholarship Repository. It has been accepted for inclusion in Florida State University Law Review by an authorized editor of Scholarship Repository. For more information, please contact [email protected]. LAW IN SOCIOBIOLOGICAL PERSPECTIVE DR. MARGARET GRUTER* I. INTRODUCTION This article is intended to acquaint legal scholars with recent findings in the biologically based behavioral sciences which may be relevant to an understanding of legal phenomena. The ideas expressed herein may stimulate further inquiries and research into the inter- action of law and behavior and may help bridge the gap between the natural sciences and the empirical studies of law. Legal research will keep pace more effectively with rapid changes in human society if the findings of the basic sciences are known and accepted by legal scholars. In view of the danger to the very continua- tion of the human species, due in part to the advancement in the natural sciences, legal research is of the utmost importance. Knowledge is neutral: what we do with it is crucial. Scientific research, with the help of law, could better our social order, but only if law builds on scientific research. -
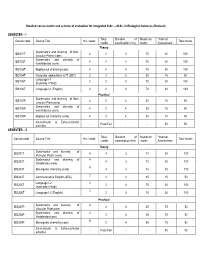
Detailed Course Matrix and Scheme of Evaluation for Integrated B.Sc. – M.Sc. in Biological Sciences (Revised) SEMESTER –
Detailed course matrix and scheme of evaluation for Integrated B.Sc. – M.Sc. in Biological Sciences (Revised) SEMESTER – I Total Duration of Maximum *Internal Course code Course Title Hrs / week Total marks credits examination (hrs) marks Assessment Theory Systematics and diversity of Non- IBS101T 4 4 3 70 30 100 vascular Plants (core) Systematics and diversity of IBS102T 4 4 3 70 30 100 Invertebrates (core) IBS103T Biophysical chemistry(core) 4 4 3 70 30 100 IBS104T Computer applications & IT (SEC) 2 2 2 35 15 50 Language I-1 IBS105T 2 2 3 70 30 100 (Kannada / Hindi) IBS106T Language I-1 (English) 2 2 3 70 30 100 Practical Systematics and diversity of Non- IBS107P 4 2 4 35 15 50 vascular Plant (core) Systematics and diversity of IBS108P 4 2 4 35 15 50 Invertebrates (core) IBS109P Biophysical chemistry (core) 4 2 4 35 15 50 Co-curricular & Extra-curricular - Pass/Fail - - 50 50 activities SEMESTER – II Total Duration of Maximum *Internal Course code Course Title Hrs / week Total marks credits examination (hrs) marks Assessment Theory Systematics and diversity of IBS201T 4 4 3 70 30 100 Vascular Plants (core) Systematics and diversity of 4 IBS202T 4 3 70 30 100 Vertebrates (core) 4 IBS203T Bio-organic chemistry (core) 4 3 70 30 100 2 IBS204T Communicative English (AEC) 2 2 35 15 50 Language I-2 2 IBS205T 2 3 70 30 100 (Kannada / Hindi) 2 IBS206T Language I-2 (English) 2 3 70 30 100 Practical Systematics and diversity of 4 IBS207P 2 4 35 15 50 Vascular Plant(core) Systematics and diversity of 4 IBS208P 2 4 35 15 50 Vertebrates(core) 4 IBS209P -

INTRODUCTION to GENETICS Table of Contents Heredity, Historical
INTRODUCTION TO GENETICS Table of Contents Heredity, historical perspectives | The Monk and his peas | Principle of segregation Dihybrid Crosses | Mutations | Genetic Terms | Links Heredity, Historical Perspective | Back to Top For much of human history people were unaware of the scientific details of how babies were conceived and how heredity worked. Clearly they were conceived, and clearly there was some hereditary connection between parents and children, but the mechanisms were not readily apparent. The Greek philosophers had a variety of ideas: Theophrastus proposed that male flowers caused female flowers to ripen; Hippocrates speculated that "seeds" were produced by various body parts and transmitted to offspring at the time of conception, and Aristotle thought that male and female semen mixed at conception. Aeschylus, in 458 BC, proposed the male as the parent, with the female as a "nurse for the young life sown within her". During the 1700s, Dutch microscopist Anton van Leeuwenhoek (1632-1723) discovered "animalcules" in the sperm of humans and other animals. Some scientists speculated they saw a "little man" (homunculus) inside each sperm. These scientists formed a school of thought known as the "spermists". They contended the only contributions of the female to the next generation were the womb in which the homunculus grew, and prenatal influences of the womb. An opposing school of thought, the ovists, believed that the future human was in the egg, and that sperm merely stimulated the growth of the egg. Ovists thought women carried eggs containing boy and girl children, and that the gender of the offspring was determined well before conception. -

The Use of Referential Gestures in Ravens (Corvus Corax) in the Wild
ARTICLE Received 5 Aug 2011 | Accepted 26 Oct 2011 | Published 29 Nov 2011 DOI: 10.1038/ncomms1567 The use of referential gestures in ravens (Corvus corax) in the wild Simone Pika1 & Thomas Bugnyar2 Around the age of one year, human children start to use gestures to coordinate attention towards a social partner and an object of mutual interest. These referential gestures have been suggested as the foundation to engage in language, and have so far only been observed in great apes. Virtually nothing is known about comparable skills in non-primate species. Here we record thirty-eight social interactions between seven raven (Corvus corax) dyads in the Northern Alps, Austria during three consecutive field seasons. All observed behaviours included the showing and/or offering of non-edible items (for example, moss, twigs) to recipients, leading to frequent orientation of receivers to the object and the signallers and subsequent affiliative interactions. We report evidence that the use of declarative gestures is not restricted to the primate lineage and that these gestures may function as ‘testing-signals’ to evaluate the interest of a potential partner or to strengthen an already existing bond. 1 Max Planck Institute for Ornithology, Research Group ‘Comparative Gestural Signalling’, Eberhard-Gwinner-Straße, Geesehouse, 82319 Seewiesen, Germany. 2 Department of Cognitive Biology, University of Vienna, 1010 Vienna, Austria. Correspondence and requests for materials should be addressed to S.P. (email: [email protected]). NatURE COMMUNicatiONS | 2:560 | DOI: 10.1038/ncomms1567 | www.nature.com/naturecommunications © 2011 Macmillan Publishers Limited. All rights reserved. ARTICLE NatUre cOMMUNicatiONS | DOI: 10.1038/ncomms1567 rom early childhood on, human infants frequently use dis- crucial in boosting not only their cognitive but also, especially, their tinct gestures such as showing, offering, giving (for example, vocal and nonvocal communicative abilities. -

Animal Umwelten in a Changing World
Tartu Semiotics Library 18 Tartu Tartu Semiotics Library 18 Animal umwelten in a changing world: Zoosemiotic perspectives represents a clear and concise review of zoosemiotics, present- ing theories, models and methods, and providing interesting examples of human–animal interactions. The reader is invited to explore the umwelten of animals in a successful attempt to retrieve the relationship of people with animals: a cornerstone of the past common evolutionary processes. The twelve chapters, which cover recent developments in zoosemiotics and much more, inspire the reader to think about the human condition and about ways to recover our lost contact with the animal world. Written in a clear, concise style, this collection of articles creates a wonderful bridge between Timo Maran, Morten Tønnessen, human and animal worlds. It represents a holistic approach Kristin Armstrong Oma, rich with suggestions for how to educate people to face the dynamic relationships with nature within the conceptual Laura Kiiroja, Riin Magnus, framework of the umwelt, providing stimulus and opportuni- Nelly Mäekivi, Silver Rattasepp, ties to develop new studies in zoosemiotics. Professor Almo Farina, CHANGING WORLD A IN UMWELTEN ANIMAL Paul Thibault, Kadri Tüür University of Urbino “Carlo Bo” This important book offers the first coherent gathering of perspectives on the way animals are communicating with each ANIMAL UMWELTEN other and with us as environmental change requires increasing adaptation. Produced by a young generation of zoosemiotics scholars engaged in international research programs at Tartu, IN A CHANGING this work introduces an exciting research field linking the biological sciences with the humanities. Its key premises are that all animals participate in a dynamic web of meanings WORLD: and signs in their own distinctive styles, and all animal spe- cies have distinctive cultures. -

2014-2015 and Is Accurate and Current, to the Greatest Extent Possible, As of June 2014
Cover Cover 1 University’s Mission Statement James B. Duke’s founding Indenture of Duke University directed the members of the University to “provide real leadership in the educational world” by choosing individuals of “outstanding character, ability and vision” to serve as its officers, trustees and faculty; by carefully selecting students of “character, determination and application;” and by pursuing those areas of teaching and scholarship that would “most help to develop our resources, increase our wisdom and promote human happiness.” To these ends, the mission of Duke University is to provide a superior liberal education to undergraduate students, attending not only to their intellectual growth but also to their development as adults committed to high ethical standards and full participation as leaders in their communities; to prepare future members of the learned professions for lives of skilled and ethical service by providing excellent graduate and professional education; to advance the frontiers of knowledge and contribute boldly to the international community of scholarship; to promote an intellectual environment built on a commitment to free and open inquiry; to help those who suffer, cure disease and promote health, through sophisticated medical research and thoughtful patient care; to provide wide ranging educational opportunities, on and beyond our campuses, for traditional students, active professionals and life-long learners using the power of information technologies; and to promote a deep appreciation for the range of human difference and potential, a sense of the obligations and rewards of citizenship, and a commitment to learning, freedom and truth. By pursuing these objectives with vision and integrity, Duke University seeks to engage the mind, elevate the spirit, and stimulate the best effort of all who are associated with the University; to contribute in diverse ways to the local community, the state, the nation and the world; and to attain and maintain a place of real leadership in all that we do. -

Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure
Loyola University Chicago Loyola eCommons Dissertations Theses and Dissertations 2018 Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure Annadorothea Asimes Loyola University Chicago Follow this and additional works at: https://ecommons.luc.edu/luc_diss Part of the Neurosciences Commons Recommended Citation Asimes, Annadorothea, "Mechanisms and Consequences of Epigenetic Inheritance Following Parental Preconception Alcohol Exposure" (2018). Dissertations. 2770. https://ecommons.luc.edu/luc_diss/2770 This Dissertation is brought to you for free and open access by the Theses and Dissertations at Loyola eCommons. It has been accepted for inclusion in Dissertations by an authorized administrator of Loyola eCommons. For more information, please contact [email protected]. Copyright © 2018 Annadorothea Asimes LOYOLA UNIVERSITY CHICAGO MECHANISMS AND CONSEQUENCES OF EPIGENETIC INHERITANCE FOLLOWING PARENTAL PRECONCEPTION ALCOHOL EXPOSURE A DISSERTATION SUBMITTED TO THE FACULTY OF THE GRADUATE SCHOOL IN CANDIDACY FOR THE DEGREE OF DOCTOR OF PHILOSOPHY PROGRAM IN NEUROSCIENCE BY ANNADOROTHEA ASIMES CHICAGO, IL MAY 2018 Copyright by AnnaDorothea Asimes, 2018 ALL rights reserved. ACKNOWLEDGEMENTS The work required to compLete this dissertation wouLd not have been possibLe alone. I wouLd first Like to thank my advisor and mentor, Dr. Toni Pak. The guidance and support you have given me throughout these graduate schooL years has been invaluabLe. You have provided a chalLenging environment that pushed me to be resourcefuL, creative, boLd, and humbLed. I wouLd not be the scientist I am today without your mentorship and friendship. I wouLd also Like to thank my Lab mates, who quickLy turned from coworkers to friends. I Look forward to buiLding our Lab famiLy and seeing the great work you alL go on to do. -

Das Egoistische Gen
Das egoistische Gen Mit einem Vorwort von Wolfgang Wickler Bearbeitet von Richard Dawkins 2., unveränderte Auflage 2014. Buch. XL, 489 S. Kartoniert ISBN 978 3 642 55390 5 Format (B x L): 12,7 x 19 cm Weitere Fachgebiete > Chemie, Biowissenschaften, Agrarwissenschaften > Tierkunde, Zoologie schnell und portofrei erhältlich bei Die Online-Fachbuchhandlung beck-shop.de ist spezialisiert auf Fachbücher, insbesondere Recht, Steuern und Wirtschaft. Im Sortiment finden Sie alle Medien (Bücher, Zeitschriften, CDs, eBooks, etc.) aller Verlage. Ergänzt wird das Programm durch Services wie Neuerscheinungsdienst oder Zusammenstellungen von Büchern zu Sonderpreisen. Der Shop führt mehr als 8 Millionen Produkte. Vorwort zur deutschen Ausgabe Die Biologie ist zur Jahrhundertwissenschaft geworden. Genetik, Mole- kularbiologie und Evolutionstheorie haben uns einen neuen Verstehens- horizont erschlossen, der gleichermaßen bedeutsam ist für Biologie wie für Medizin und Philosophie, für das Verständnis und den Umgang mit der Natur wie für Schöpfungsvorstellungen. Dazu hat Richard Dawkins eine entscheidende neue Idee hinreißend formuliert. Sein sachlich über- raschendes, genial gedachtes und rasant geschriebenes Buch hat ihm je- doch nicht nur Beifalls-, sondern auch Entrüstungsstürme eingebracht. Ein Grund für letztere liegt wohl darin, daß er einen Paradigmenwechsel für Laien verständlich erläuterte, noch bevor viele Fachleute ihn begrif- fen hatten. Dawkins tut das, indem er die Theorien mit markanten Bei- spielen illustriert und nicht umständlich versucht, ihre Richtigkeit zu be- weisen. Worum geht es? Noch zu Darwins Lebzeiten hatte sich in europäische Denkgewohnheiten die Idee der Arterhaltung fest eingenistet. Es schien das Natürlichste von der Welt, daß alle Lebewesen danach strebten, ihre Art zu erhalten. Schon im 13. Jahrhundert hatte Thomas von Aquin die Arterhaltung zu den grundlegenden natürlichen Neigungen gezählt.