Functional Morphology of the Distal Forelimb and the Evolution of Tool Use in Humans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
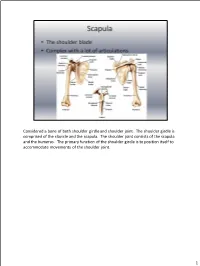
Considered a Bone of Both Shoulder Girdle and Shoulder Joint. the Shoulder Girdle Is Comprised of the Clavicle and the Scapula
Considered a bone of both shoulder girdle and shoulder joint. The shoulder girdle is comprised of the clavicle and the scapula. The shoulder joint consists of the scapula and the humerus. The primary function of the shoulder girdle is to position itself to accommodate movements of the shoulder joint. 1 Superior angle—top point Inferior angle—bottom point Vertebral border—side closest to vertebral column Axillary border—side closest to arm Subscapular fossa—anterior fossa Glenoid fossa, glenoid labrum, glenoid cavity --The glenoid fossa is the shallow cavity where the humeral head goes. The glenoid labrum is the cartilage that goes around the glenoid fossa. So the glenoid fossa and glenoid labrum together comprise the glenoid cavity. Supraspinous fossa—posterior, fossa above the spine Spine of the scapula—the back projection Infraspinous fossa—posterior depression/fossa below spine Coracoid process—anterior projection head Acromion process—posterior projection head above spine 2 Scapulothoracic “joint” = NOT a true joint; there are no ligaments or articular capsule. The scapula just rests on the muscle over top the rib cage, which allows for passive movements. Sternoclavicular joint=where the clavicle (collarbone) and the sternum (breastbone) articulate; movement is slight in all directions and of a gliding, rotational type Acromioclavicular joint = where the clavicle and scapula (acromion process) articulate; AKA: AC Joint; movement is a slight gliding when elevation and depression take place. Glenohumeral joint = the shoulder joint 3 4 All 3 true joints: Sternoclavicular, AC and glenohumeral (GH) all work together to move arm in all directions. The GH allows the arm to go out to the side and be abducted, then the AC and Sternoclavicular joints kick in to allow the arm to go above shoulder level by allowing the shoulderblade to move up to increase the range of motion (ROM). -

The Appendicular Skeleton Appendicular Skeleton
THE SKELETAL SYSTEM: THE APPENDICULAR SKELETON APPENDICULAR SKELETON The primary function is movement It includes bones of the upper and lower limbs Girdles attach the limbs to the axial skeleton SKELETON OF THE UPPER LIMB Each upper limb has 32 bones Two separate regions 1. The pectoral (shoulder) girdle (2 bones) 2. The free part (30 bones) THE PECTORAL (OR SHOULDER) GIRDLE UPPER LIMB The pectoral girdle consists of two bones, the scapula and the clavicle The free part has 30 bones 1 humerus (arm) 1 ulna (forearm) 1 radius (forearm) 8 carpals (wrist) 19 metacarpal and phalanges (hand) PECTORAL GIRDLE - CLAVICLE The clavicle is “S” shaped The medial end articulates with the manubrium of the sternum forming the sternoclavicular joint The lateral end articulates with the acromion forming the acromioclavicular joint THE CLAVICLE PECTORAL GIRDLE - CLAVICLE The clavicle is convex in shape anteriorly near the sternal junction The clavicle is concave anteriorly on its lateral edge near the acromion CLINICAL CONNECTION - FRACTURED CLAVICLE A fall on an outstretched arm (F.O.O.S.H.) injury can lead to a fractured clavicle The clavicle is weakest at the junction of the two curves Forces are generated through the upper limb to the trunk during a fall Therefore, most breaks occur approximately in the middle of the clavicle PECTORAL GIRDLE - SCAPULA Also called the shoulder blade Triangular in shape Most notable features include the spine, acromion, coracoid process and the glenoid cavity FEATURES ON THE SCAPULA Spine - -

Bone Limb Upper
Shoulder Pectoral girdle (shoulder girdle) Scapula Acromioclavicular joint proximal end of Humerus Clavicle Sternoclavicular joint Bone: Upper limb - 1 Scapula Coracoid proc. 3 angles Superior Inferior Lateral 3 borders Lateral angle Medial Lateral Superior 2 surfaces 3 processes Posterior view: Acromion Right Scapula Spine Coracoid Bone: Upper limb - 2 Scapula 2 surfaces: Costal (Anterior), Posterior Posterior view: Costal (Anterior) view: Right Scapula Right Scapula Bone: Upper limb - 3 Scapula Glenoid cavity: Glenohumeral joint Lateral view: Infraglenoid tubercle Right Scapula Supraglenoid tubercle posterior anterior Bone: Upper limb - 4 Scapula Supraglenoid tubercle: long head of biceps Anterior view: brachii Right Scapula Bone: Upper limb - 5 Scapula Infraglenoid tubercle: long head of triceps brachii Anterior view: Right Scapula (with biceps brachii removed) Bone: Upper limb - 6 Posterior surface of Scapula, Right Acromion; Spine; Spinoglenoid notch Suprspinatous fossa, Infraspinatous fossa Bone: Upper limb - 7 Costal (Anterior) surface of Scapula, Right Subscapular fossa: Shallow concave surface for subscapularis Bone: Upper limb - 8 Superior border Coracoid process Suprascapular notch Suprascapular nerve Posterior view: Right Scapula Bone: Upper limb - 9 Acromial Clavicle end Sternal end S-shaped Acromial end: smaller, oval facet Sternal end: larger,quadrangular facet, with manubrium, 1st rib Conoid tubercle Trapezoid line Right Clavicle Bone: Upper limb - 10 Clavicle Conoid tubercle: inferior -

Shoulder Shoulder
SHOULDER SHOULDER ⦿ Connects arm to thorax ⦿ 3 joints ◼ Glenohumeral joint ◼ Acromioclavicular joint ◼ Sternoclavicular joint ⦿ https://www.youtube.com/watch?v=rRIz6oO A0Vs ⦿ Functional Areas ◼ scapulothoracic ◼ scapulohumeral SHOULDER MOVEMENTS ⦿ Global Shoulder ⦿ Arm (Shoulder Movement Joint) ◼ Elevation ◼ Flexion ◼ Depression ◼ Extension ◼ Abduction ◼ Abduction ◼ Adduction ◼ Adduction ◼ Medial Rotation ◼ Medial Rotation ◼ Lateral Rotation ◼ Lateral Rotation SHOULDER MOVEMENTS ⦿ Movement of shoulder can affect spine and rib cage ◼ Flexion of arm Extension of spine ◼ Extension of arm Flexion of spine ◼ Adduction of arm Ipsilateral sidebending of spine ◼ Abduction of arm Contralateral sidebending of spine ◼ Medial rotation of arm Rotation of spine ◼ Lateral rotation of arm Rotation of spine SHOULDER GIRDLE ⦿ Scapulae ⦿ Clavicles ⦿ Sternum ⦿ Provides mobile base for movement of arms CLAVICLE ⦿ Collarbone ⦿ Elongated S shaped bone ⦿ Articulates with Sternum through Manubrium ⦿ Articulates with Scapula through Acromion STERNOCLAVICULAR JOINT STERNOCLAVICULAR JOINT ⦿ Saddle Joint ◼ Between Manubrium and Clavicle ⦿ Movement ◼ Flexion - move forward ◼ Extension - move backward ◼ Elevation - move upward ◼ Depression - move downward ◼ Rotation ⦿ Usually movement happens with scapula Scapula Scapula ● Flat triangular bone ● 3 borders ○ Superior, Medial, Lateral ● 3 angles ○ Superior, Inferior, Lateral ● Processes and Spine ○ Acromion Process, Coracoid Process, Spine of Scapula ● Fossa ○ Supraspinous, Infraspinous, Subscapularis, Glenoid SCAPULA -

The Ulna Humerus and Ulna: Landmarks and Articulations
This document was created by Alex Yartsev ([email protected]); if I have used your data or images and forgot to reference you, please email me. The Ulna o the ulna is a medial long bone, the longer of the two in the forearm. Proximally, it articulates with the capitulum and trochlea of the humerus; . At the radial notch, it articulates with the head of radius o It stabilizes the forearm o The HEAD LIES DISTALLY. The Olecranon The Trochlear Notch The Coronoid Process The Radial Notch where the head of the radius goes The Tuberosity of the Ulna is where the Brachialis attaches The deep part of Supinator fossa the supinator Supinator crest attaches here Bony features include: - Olecranon - Trochlear notch - Coronoid process - Radial notch - Tuberosity of ulna - Supinator fosa - Supinator crest - Head of ulna - Ulnar styloid process The head of ulna The ulnar styloid process Humerus and ulna: landmarks and articulations o the humerus is a long bone, the largest in the upper limb o it articulates proximally with the scapula at the scapulohumeral (glenohumeral) joint o it articulates distally with the ulna at the elbow joint o the ulna and humerus articulate at the elbow joint o the articulations include: . articulation between the trochlea of the humerus and the trochlear notch of the ulna . articulation of the olecranon process and the olecranon fossa during extension . articulation of the coronoid process and the coronoid fossa during flexion o the surface landmarks include . the medial and lateral epicondyle . the olecranon . the posterior border of the ulna . -
Lab Manual Appendicular Skele
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the appendicular skeleton, which is formed from the pectoral and pelvic girdles and the upper and lower limbs. View Module 7.2 Axial and Appendicular Skeleton to highlight the bones of the appendicular skeleton and compare them to those of the axial skeleton. Examine Module 11.1 Appendicular Skeleton to view only the bones of the appendicular skeleton. In addition to learning about all the bones of the appendicular skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Proximal: Trochanter: Tubercle: Tuberosity: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the appendicular skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. -

Olecranon Fractures Injury
Injury, Int. J. Care Injured 40 (2009) 575–581 Contents lists available at ScienceDirect Injury journal homepage: www.elsevier.com/locate/injury Review Olecranon fractures S.D.S. Newman a,*, C. Mauffrey b, S. Krikler c a Milton Keynes General Hospital, UK b Birmingham Heartlands and Solihull Hospital, UK c University Hospital Coventry and Warwickshire, UK ARTICLE INFO ABSTRACT Article history: Several options exist for the management olecranon fractures. These include tension band, plate and Accepted 9 December 2008 intramedullary fixation techniques as well as fragment excision with triceps advancement and non- operative management. No one technique is suitable for the management of all olecranon fractures. In Keywords: deciding how to treat this common trauma presentation, the surgeon needs a good understanding of the Olecranon fracture anatomy, different fracture morphologies, surgical options and potential complications. With Tension band fixation appropriate management and early mobilisation good functional results can be expected in the Plate fixation majority of patients. Intramedullary fixation Crown Copyright ß 2008 Published by Elsevier Ltd. All rights reserved. Elbow anatomy Fracture classification Contents Introduction . .................................................................................... 575 Anatomy ........................................................................................ 576 Mechanism of injury ................................................................................ 576 Classification . ................................................................................... -

Ulnar Trochlear Notch Articular Surface Has Three Morphological Patterns: a Neglected Major Anatomical Feature
Surgical and Radiologic Anatomy (2019) 41:1333–1336 https://doi.org/10.1007/s00276-019-02310-y ORIGINAL ARTICLE Ulnar trochlear notch articular surface has three morphological patterns: a neglected major anatomical feature Trifon Totlis1 · Nikolaos Otountzidis1 · Stavros Papadopoulos1 · Maria Piagkou2 · Konstantinos Natsis1 Received: 13 March 2019 / Accepted: 23 August 2019 / Published online: 31 August 2019 © Springer-Verlag France SAS, part of Springer Nature 2019 Abstract Purpose The typical anatomical illustration of the trochlear notch articular surface includes a transverse rough non-articular ridge which separates the olecranon and coronoid part of the joint. This transverse ridge, the “bare area”, is not covered by cartilage and represents the optimal entry point for olecranon osteotomies. Aim of the present study was to encounter the anatomical variations in the morphology of the trochlear notch articular surface. Methods Two-hundred seventy-three dried ulnae were inspected and a qualitative classifcation of the variations of the trochlear notch articular surface was done. Correlation to gender and side was examined. Results Three distinct morphological patterns were observed. Separate olecranon and coronoid facets (Type I) were the most common pattern (165/273, 60.4%). Partial fusion of olecranon and coronoid facets (Type II) was observed in 75 out of 273 bones (27.5%), while a single olecranon and coronoid facet (Type III) was present in 33 out of 273 bones (12.1%). The occurrence of Type II and III was signifcantly more common in females (p < 0.001). Conclusions The most common morphological pattern of the proximal ulna articular surface was the olecranon and coro- noid facets separated by the transverse ridge, which is considered as the typical anatomical pattern. -

An Anthropometric Investigation Into the Probable Cause of Formation of 'Carrying Angle': a Sex Indicator
JIAFM, 2004; 26(1). ISSN 0971-0973 AN ANTHROPOMETRIC INVESTIGATION INTO THE PROBABLE CAUSE OF FORMATION OF ‘CARRYING ANGLE’: A SEX INDICATOR Dr. Ruma Purkait, Senior Lecturer, Department of Anthropology, Saugor University, Saugor Dr. Heeresh Chandra, Founder & Former Director, Medicolegal Institute, Bhopal, M.P ABSTRACT In the living the ‘Carrying angle’ measures around 173 degrees in males and 167 degrees in females. The cause of its formation is a long debated issue. The present study is an attempt to identify by anthropometric means the sexually dimorphic features in the bones of the elbow joint which makes the ‘Carrying angle’ a sex indicator. The distal end of the humerus and the proximal end of ulna playing major role in the formation of ‘Carrying angle’ have been examined for sex difference. The two measurements of the humerus (Trochlear angle and Inclination angle of Olecranon fossa) and three measurements of the ulna (Olecranon – coronoid angle, length and width of inferior medial trochlear notch) were devised for the study. Though the humeral angles failed to show any sex difference, the angle and dimensions of ulna exhibited statistically significant result. Could these parameters be the cause of sex differences the ‘Carrying angle’ exhibits at the elbow joint? KEY WORDS: ‘Carrying angle’, sexual dimorphism, humerus, ulna INTRODUCTION debated issue in two ‘A’s, Anatomy and When the forearm is completely extended Anthropology. Surveying the available literature on and supinated, the long axis beyond the elbow joint the subject indicates that authors have expressed is not in line with the upper arm but is deviated different views about its cause of formation. -
Stress Distribution in the Trochlear Notch
STRESS DISTRIBUTION IN THE TROCHLEAR NOTCH A MODEL OF BICENTRIC LOAD TRANSMISSION ThROUGH JOINTS F. ECKSTEIN, F. LOHE, M. MULLER-GERBL, M. STEINLECHNER, R. PUTL From Ludwig-Maximilians-Universizy ofMunich, Germany In 16 cadaver humeroulnar joints, the distribution because the distribution of pressure within the contact of subchondral mineralisation was assessed by CT zones may not be uniform (Hehne 1983; Miyanaga et al osteoabsorptiometry and the position and size of the 1984). contact areas by polyether casting under loads of 10 N It is possible, however, to assess the physiologically to 1280 N. effective pressure distribution over the surface of a joint Ulnas with separate olecranon and coronoid from the functional adaptation of the subchondral tissues cartilaginous surfaces showed matching bicentric (Kummer 1962; Tillmann 1978; Pauwels 1980). Using patterns of mineralisation. Under small loads there the theory of ‘causal histogenesis’, M#{252}ller-Gerbl and her were separate contact areas on the olecranon and co-workers (1989, 1990, 1992) have developed the coronoid surfaces; these areas merged centrally as the technique of CT osteoabsorptiometry (CT-OAM) to load increased. They occupied as little as 9% of the produce a surface projection of the distribution of total articular surface at 10 N and up to 73 % at subchondral mineralisation. The density pattern obtained 1280 N. Ulnas with continuous cartilaginous surfaces with this method reflects the ‘loading history’ (Carter also had density patterns with two maxima but those 1984; Carter, Wong and Orr 1991) of the joint surface were less prominent, and in these specimens the during daily life. It is an expression of the sum of a vast separate contact areas merged at lower loads. -
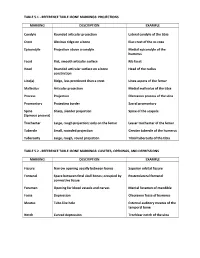
Table 5.1 - Reference Table: Bone Markings: Projections
TABLE 5.1 - REFERENCE TABLE: BONE MARKINGS: PROJECTIONS MARKING DESCRIPTION EXAMPLE Condyle Rounded articular projection Lateral condyle of the tibia Crest Obvious ridge on a bone Iliac crest of the os coxa Epicondyle Projection above a condyle Medial epicondyle of the humerus Facet Flat, smooth articular surface Rib facet Head Rounded articular surface on a bone Head of the radius constriction Line(a) Ridge, less prominent than a crest Linea aspera of the femur Malleolus Articular projection Medial malleolus of the tibia Process Projection Olecranon process of the ulna Promontory Projecting border Sacral promontory Spine Sharp, slender projection Spine of the scapula (Spinous process) Trochanter Large, rough projection; only on the femur Lesser trochanter of the femur Tubercle Small, rounded projection Greater tubercle of the humerus Tuberosity Large, rough, round projection Tibial tuberosity of the tibia TABLE 5.2 - REFERENCE TABLE: BONE MARKINGS: CAVITIES, OPENINGS, AND DEPRESSIONS MARKING DESCRIPTION EXAMPLE Fissure Narrow opening usually between bones Superior orbital fissure Fontanel Space between fetal skull bones; occupied by Posterolateral fontanel connective tissue Foramen Opening for blood vessels and nerves Mental foramen of mandible Fossa Depression Olecranon fossa of humerus Meatus Tube-like hole External auditory meatus of the temporal bone Notch Curved depression Trochlear notch of the ulna TABLE 5.3 - AXIAL SKELETON: SKULL BONES – CRANIAL BONES BONE MARKINGS Frontal Parietal Occipital Foramen magnum, Occipital condyles -

Complete Thesis
2. Review Literature 2.1 Anatomy and development of the elbow joint The elbow joint (cubital joint) is a stable compound ginglymus or hinge joint: It is composed of the humero-radial joint formed by the humeral condyle and head of the radius, the humero-ulnar joint, which is formed by the semilunar notch of the ulna and humeral condyle, and the proximal radio-ulnar joint (FOX et al., 1983; LEWIS et al., 1989; NICKEL et al., 1992; EVANS, 1993; VOLLMERHAUS et al., 1994). The proximal radio-ulnar joint is composed of the articular circumference of the radial head with the radial notch of the ulna (LEWIS et al., 1989). All components share a common joint capsule. The elbow joint enables flexion, extension and a limited amount of rotation. The humero-radial joint is responsible for 75% to 80% of weight-bearing surface of the joint (BERZON and QUICK, 1980) while the humero-ulnar part stabilizes and restricts the movement of the joint in the sagittal plane and the proximal radio-ulnar joint allows rotation of the antebrachium (BERZON and QUICK, 1980; FOX et al., 1983; EVANS, 1993) The anconeal process of the ulna articulates with the caudal intercondylar surface of the humerus and fits into the supratrochlear fossa when the joint is fully extended. The trochlear notch of the ulna articulates with the trochlear of the humerus. Distal to the trochlear notch are two prominences called the medial and lateral coronoid processes. The medial process is larger and is located more distal than the lateral (BERZON and QUICK, 1980; FOX et al., 1983).