The Key Role of Patrilineal Inheritance in Shaping the Genetic Variation of Dagestan Highlanders
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Humanitarian Action in the Caucasus: a Guide for Practitioners
HUMANITARIAN ACTION IN THE CAUCASUS: A GUIDE FOR PRACTITIONERS Greg Hansen Humanitarianism and War Project & Local Capacities for Peace Project i Published by The Thomas J. Watson Jr. Institute for International Studies Brown University, Box 1970 2 Stimson Avenue Providence, RI 02912 USA Telephone: (401) 863-2809 Fax: (401) 863-1270 E-mail: [email protected] http://www.brown.edu/Departments/Watson_Institute/ Thomas J. Biersteker, Ph.D., Director Frederick F. Fullerton, Writer/Editor Nancy Soukup, Writer/Editor George Potter, Staff Assistant Statements of fact or opinions are solely those of the authors; their publication does not imply endorsement by the Thomas J. Watson Jr. Institute for International Studies. Copyright 1998 by the Thomas J. Watson Jr. Institute for International Studies. All rights reserved under International and Pan American Convention. No part of this report may be reproduced by any other means, electronic or mechanical, including photocopy, recording, or any information storage and retrieval system, without prior written permission from the publisher. All inquiries should be addressed to Publications Group, Thomas J. Watson Jr. Institute for International Studies. ii CONTENTS Preface ........................................................................................................................................... v Author’s Note...............................................................................................................................ix Acronyms......................................................................................................................................xi -

World Directory of Minorities
World Directory of Minorities Europe MRG Directory –> Russian Federation –> Avars Print Page Close Window Avars Profile According to the 2002 national census, there are 814,473 Avars in the Russian Federation. Avars are a mountain people and are numerically the largest group in Dagestan. They account for approximately 28 per cent of the population of Dagestan and are one of the republic's 14 official ‘titular' nationalities. Small populations of Avars also reside in Georgia and Azerbaijan. Historical context The Avar people were constituted as a singular ‘official' nationality from a large variety of culturally related local groups in the 1930s. Avars in Dagestan inhabit primarily mono-ethnic districts. Traditionally Avars have played a pre-eminent role in the delicate, informal power-sharing system between the many ethnic groups in Dagestan. This system has successfully contained a number of traditional rivalries, for instance, that between Caucasian highlanders, such as the Avars, and Turkic lowlanders. An example is the repeated rejection in three post-Soviet referenda in Dagestan that would have established a presidency, an institution that would concentrate power in the hands of one ethnic group over others. Avars played a central role in blocking the introduction of a presidency, which would have diminished their numerical advantage. Instead, Dagestan's 1993 constitution provides for a collective presidency, known as the State Council, composed of 14 members, one from each of Dagestan's 14 official titular nationalities - including Avars. Although the chairmanship of the State Council was originally intended to be a post rotating between ethnic groups, it was abolished in 1998 in recognition of the de facto control over the chairmanship of an ethnic Dargin. -

A Balkanist in Daghestan: Annotated Notes from the Field Victor A
A Balkanist in Daghestan: Annotated Notes from the Field Victor a. Friedman University of Chicago Introduction and Disclaimer The Republic of Daghestan has received very little attention in the West. Chenciner (1997) is the only full-length account in English based on first-hand visits mostly in the late 1980's and early 1990's. Wixman's (1980) excellent study had to be based entirely on secondary sources, and Bennigsen and Wimbush (1986:146-81 et passim), while quite useful, is basically encyclopedic and somewhat dated. Since Daghestan is still difficult to get to, potentially unstable, and only infrequently visited by Western scholars (mostly linguists), I am offering this account of my recent visit there (16-20 June 1998), modestly supplemented by some published materials. My intent is basically informative and impressionistic, and I do not attempt to give complete coverage to many topics worthy of further research. This account does, however, update some items covered in the aforementioned works and makes some observations on Daghestan with respect to language, identity, the political situation, and a comparison with the another unstable, multi-ethnic, identity construction site, i.e., Balkans, particularly Macedonia. Background Daghestan is the third most populous Republic in the Russian Federation (after Bashkortostan and Tatarstan; Osmanov 1986:24). The northern half of its current territory, consisting of the Nogai steppe and the Kizljar region settled in part by Terek Cossacks, was added in 1922, after the fall of the North Caucasian -

Russia's Peacetime Demographic Crisis
the national bureau of asian research nbr project report | may 2010 russia’s peacetime demographic crisis: Dimensions, Causes, Implications By Nicholas Eberstadt ++ The NBR Project Report provides access to current research on special topics conducted by the world’s leading experts in Asian affairs. The views expressed in these reports are those of the authors and do not necessarily reflect the views of other NBR research associates or institutions that support NBR. The National Bureau of Asian Research is a nonprofit, nonpartisan research institution dedicated to informing and strengthening policy. NBR conducts advanced independent research on strategic, political, economic, globalization, health, and energy issues affecting U.S. relations with Asia. Drawing upon an extensive network of the world’s leading specialists and leveraging the latest technology, NBR bridges the academic, business, and policy arenas. The institution disseminates its research through briefings, publications, conferences, Congressional testimony, and email forums, and by collaborating with leading institutions worldwide. NBR also provides exceptional internship opportunities to graduate and undergraduate students for the purpose of attracting and training the next generation of Asia specialists. NBR was started in 1989 with a major grant from the Henry M. Jackson Foundation. Funding for NBR’s research and publications comes from foundations, corporations, individuals, the U.S. government, and from NBR itself. NBR does not conduct proprietary or classified research. The organization undertakes contract work for government and private-sector organizations only when NBR can maintain the right to publish findings from such work. To download issues of the NBR publications, please visit the NBR website http://www.nbr.org. -

State Report Azerbaijan
ACFC/SR(2002)001 ______ REPORT SUBMITTED BY AZERBAIJAN PURSUANT TO ARTICLE 25, PARAGRAPH 1 OF THE FRAMEWORK CONVENTION FOR THE PROTECTION OF NATIONAL MINORITIES ______ (Received on 4 June 2002) _____ TABLE OF CONTENTS PART I............................................................................................................................................ 3 II. Aggression of the Republic of Armenia against the Republic of Azerbaijan..................... 9 III. Information on the form of the State structure.................................................................. 12 IV. Information on status of international law in national legislation .................................... 13 V. Information on demographic situation in the country ...................................................... 13 VI. Main economic data - gross domestic product and per capita income ............................. 15 VII. State’s national policy in the field of the protection of the rights of persons belonging to minorities ...................................................................................................................................... 15 VIII. Population awareness on international treaties to which Azerbaijan is a party to........ 16 P A R T II..................................................................................................................................... 18 Article 1 ........................................................................................................................................ 18 Article -

North Caucasus: Views from Within People’S Perspectives on Peace and Security
REPORT North Caucasus: views from within People’s perspectives on peace and security March 2012 North Caucasus: views from within People’s perspectives on peace and security SAFERWORLD MARCH 2012 Acknowledgements This report was written by Anna Matveeva, Honorary University Fellow, University of Exeter, with contributions from Igor Savin, Research Fellow, the Institute of Oriental Studies, Russian Academy of Sciences. It was edited by Craig Oliphant (Senior Advisor, Saferworld). Editorial contributions and comments were also provided by Luitgard Hammerer (Head of Europe and Central Asia, Saferworld). The report draws on participatory research conducted in five North Caucasus republics by Zhanna Khamdokhova (Kabardino-Balkaria), Igor Dulaev (North Ossetia), Musa Yusupov (Chechnya), Akhmet Yarlykapov (Dagestan), Anna Matveeva and Igor Savin (Ingushetia). The methodology for the research was developed by Anna Matveeva and Igor Savin with contributions from the Institute of Oriental Studies, Saferworld and the four researchers from the North Caucasus. For further reading, case studies from the individual republics can be accessed on the Saferworld website: www.saferworld.org.uk/PPP/chechnya www.saferworld.org.uk/PPP/northossetia www.saferworld.org.uk/PPP/kabardino-balkaria www.saferworld.org.uk/PPP/ingushetia www.saferworld.org.uk/PPP/dagestan The republic chapters were translated from Russian into English by Sophia Matveeva. John Newman copy-edited the report. The research was a collaboration between Saferworld and the Institute of Oriental Studies of the Russian Academy of Sciences. The People’s Peacemaking Perspectives project The People’s Peacemaking Perspectives project is a joint initiative implemented by Conciliation Resources and Saferworld and financed under the European Commission’s Instrument for Stability. -
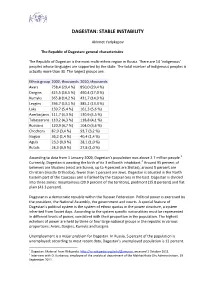
Dagestan: Stable Instability
DAGESTAN: STABLE INSTABILITY Akhmet Yarlykapov The Republic of Dagestan: general characteristics The Republic of Dagestan is the most multi-ethnic region in Russia. There are 14 ‘indigenous’ peoples whose languages are supported by the state. The total number of indigenous peoples is actually more than 30. The largest groups are: Ethnic group 2002, thousands 2010, thousands Avars 758,4 (29,4 %) 850,0 (29,4 %) Dargins 425,5 (16,5 %) 490,4 (17,0 %) Kumyks 365,8 (14,2 %) 431,7 (14,9 %) Lezgins 336,7 (13,1 %) 385,2 (13,3 %) Laks 139,7 (5,4 %) 161,3 (5,6 %) Azerbaijanis 111,7 (4,3 %) 130,9 (4,5 %) Tabassarans 110,2 (4,3 %) 118,8 (4,1 %) Russians 120,9 (4,7 %) 104,0 (3,6 %) Chechens 87,9 (3,4 %) 93,7 (3,2 %) Nogais 36,2 (1,4 %) 40,4 (1,4 %) Aguls 23,3 (0,9 %) 28,1 (1,0 %) Rutuls 24,3 (0,9 %) 27,8 (1,0 %) According to data from 1 January 2009, Dagestan’s population was above 2.7 million people. 1 Currently, Dagestan is awaiting the birth of its 3 millionth inhabitant. 2 Around 95 percent of believers are Muslims (most are Sunnis, up to 4 percent are Shiites), around 5 percent are Christian (mostly Orthodox), fewer than 1 percent are Jews. Dagestan is situated in the North Eastern part of the Caucasus and is flanked by the Caspian Sea in the East. Dagestan is divided into three zones: mountainous (39.9 percent of the territory), piedmont (15.8 percent) and flat plain (43.3 percent). -

Russia's Dagestan: Conflict Causes
RUSSIA’S DAGESTAN: CONFLICT CAUSES Europe Report N°192 – 3 June 2008 TABLE OF CONTENTS EXECUTIVE SUMMARY ...................................................................................................... i I. INTRODUCTION ............................................................................................................. 1 II. A FRAGILE INTER-ETHNIC BALANCE.................................................................... 2 A. INTER-ETHNIC COMPETITION OVER LAND AND STATE POSITIONS...............................................2 B. THE 2007 ELECTIONS .................................................................................................................4 1. Removing inter-ethnic competition from electoral politics..................................................4 2. Electoral violence and results ...............................................................................................5 III. ISLAMISM IN DAGESTAN AND CHECHEN CONNECTIONS.............................. 6 A. CHECHEN AND DAGESTANI ISLAMISTS IN THE 1990S .................................................................6 B. THE “HUNT FOR THE WAHHABIS” SINCE 1999 ...........................................................................8 C. SHARIAT JAMAAT’S GROWING INFLUENCE .................................................................................8 D. RENEWED TENSIONS WITH CHECHNYA .....................................................................................10 IV. VIOLENCE AGAINST STATE AUTHORITIES ...................................................... -

Conflicts in the North Caucasus
Central Asian Survey, (1998), 17(3), 409-441. Conflicts in the North Caucasus SVANTE E. CORNELL* Although the conflicts that have attracted most international attention in the post- cold war era have been those of the Transcaucasus, another area of both potential and actual turmoil is the North Caucasus. The first example of serious conflicts in the area, naturally, is the war in Chechnia. However, the existence of this war, and its astonishing cruelty and devastation, has been instrumental in obscuring the other grievances that exist in the area, and that have the potential to escalate into open conflict. These can be divided into two main categories, which however spill over into one another. The first type of conflicts are those among the peoples of the region; the second type is conflicts between these peoples and Russia. Doubtlessly, the main example of the unrest in the North Caucasus has been the short but bloody war between the Ingush and the Ossetians. However, this war distinguishes itself only by being the only one of the conflicts in the region that has escalated into war. Among the other known problems, three issues call for special attention: First and perhaps most pressingly, the bid for unity of the Lezgian people in Dagestan and Azerbaijan; Second, the latent problem between the Turkic Karachai and the Circasssian peoples in the two neighbour republics of Karachaevo-Cherkessia and Kabardino-Balkaria; and thirdly the condition of the most complex of all the North Caucasian republics: Dagestan. It seems logical in this study to start with the most well-known of these issues: the war that raged in November 1992 in the Prigorodniy raion between the Ingush and the North Ossetians. -

National Minorities in South Caucasus
Dr. Ahmad Shahidov Azerbaijan Institute for Democracy and Human Rights (AIDHR) www.aidhr.org [email protected] +99450 372 87 30 NATIONAL MINORITIES IN SOUTH CAUCASUS Today, Azerbaijan is distinguished with ethnic diversity. Besides Azerbaijani Turks, Mountain Jews, Tats, Talysh, Kurds, Molokans, Ingiloys, Tsakhurs, Avars, Lezgins, Khynalygs, Buduqlus, Grysz and other ethnic groups live in its territory. Although the representatives of ethnic groups consider themselves as Azerbaijanis but each group has retained distinctive elements of their different culture. This culture is reflected in domestic life, crafts, in kitchen and various ceremonies. Despite the difficulties being experienced by Azerbaijan, due to the unresolved armed conflict that continues more than a decade with neighboring Armenia, which occupies 20 percent of the Azerbaijani territory and the presence of about one million refugees and forcibly displaced persons - victims of ethnic cleansing organized by the Armenian armed forces and terrorist groups, among whom in addition to Azerbaijanis there are persons belonging to various minorities (Kurds, Russians, Jews and others), as well as problems connected with transitional period, the Government of Azerbaijan continues to pursue a consistent policy towards protection of minorities rights. 1 National minorities constitute 9.4 % of the population of the Republic of Azerbaijan. Here is a structural composition of population, languages and places of compact living of persons belonging to national minorities of the Republic of Azerbaijan: Lezgins - 178 thousand, compactly live in the Northern regions of Azerbaijan. Language of communication is Lezgins, relating to the Daghestan branch of the Caucasian languages, as well as Azerbaijani and Russian languages. Russians - 141,7 thousand, compactly live in the industrial cities, as well as a number of rural regions. -

Draft—August 1995
“RUSSIA’S TINDERBOX” Conflict in the North Caucasus And its Implications for The Future of the Russian Federation Fiona Hill September 1995 TABLE OF CONTENTS Preface Background to the Report Introduction Executive Summary Section I The Structural Legacy of the USSR Section II The Crisis of National and Regional Leadership in the North Caucasus Section III The Failure of Russia’s Regional Policy Section IV Territorial Disputes in the North Caucasus Section V Chechnya Appendix 1 The Refugee Crisis in the North Caucasus Appendix 2 The Islamic Factor in the North Caucasus Bibliography Selected Works in English for Additional Reading on the North Caucasus PREFACE Background to the Report In the Summer of 1993, the Strengthening Democratic Institutions Project began an inquiry into the conflicts in Russia’s North Caucasus region, as part of a broader study of post-Soviet ethno- political conflicts. In the course of this inquiry, it became apparent that there were practically no contemporary English-language studies of the North Caucasus. Although the new Transcaucasian states of Georgia, Armenia and Azerbaijan were beginning to be studied more closely as a consequence of their new status as independent states after 1991, information on the Russian North Caucasus region was only available from news wires and the occasional flying visits of Western correspondents. This was in spite of the prevalence of violent conflict in the Caucasus as a whole, the involvement of North Caucasian groups in these conflicts, and Chechnya’s 1991 declaration of independence from the Russian Federation. By 1993, Azerbaijan and Armenia had been in a de facto state of war over Nagorno- Karabakh for almost five years; an armed conflict had flared between Georgia and South Ossetia sending a wave of refugees into North Ossetia in the North Caucasus; North Caucasian mercenaries were fighting on the side of Abkhazia in its war with Georgia; and a violent conflict had erupted within the North Caucasus itself between Ossetians and Ingush. -

Ethnic Competition, Radical Islam, and Challenges to Stability in the Republic of Dagestan
Communist and Post-Communist Studies 43 (2010) 297–308 Contents lists available at ScienceDirect Communist and Post-Communist Studies journal homepage: www.elsevier.com/locate/postcomstud Ethnic competition, radical Islam, and challenges to stability in the Republic of Dagestan Edward C. Holland a,*, John O’Loughlin b a Institute of Behavioral Science and Department of Geography, University of Colorado, Campus Box 487, Boulder, CO 80309-0487, United States b Institute of Behavioral Science, University of Colorado, United States article info abstract Article history: Previous academic work on stability in Dagestan has focused on two potential cleavages, Available online 23 August 2010 the republic’s ethnic diversity and the challenge from radical Islamist groups. Using results from a December 2005 survey, and focusing on Dagestan’s six main ethnic groups, this paper investigates attitudes towards the dual topics of the politicization of ethnicity and Keywords: the relationship between terrorism and Islamism. We find that Dagestanis maintain Dagestan layered conceptions of identity, and do not attribute violence predominantly to radical North Caucasus Islam in the republic or the wider North Caucasus. Scholars should be aware of Rogers Nationalities ’ Radical Islam Brubaker s concept of groupism in analyzing not just ethnic groups, but religious move- Groupism ments as well. Public opinion survey Ó 2010 Published by Elsevier Ltd on behalf of The Regents of the University of California. Introduction Research on Dagestan in the post-Soviet period has noted that the republican leadership has had to “walk a tightrope between nationalism and Islam” (Gammer, 2002: p. 139) in order to maintain political stability.