Microrna in Control of Gene Expression: an Overview of Nuclear Functions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Comparison of the Effects on Mrna and Mirna Stability Arian Aryani and Bernd Denecke*
Aryani and Denecke BMC Research Notes (2015) 8:164 DOI 10.1186/s13104-015-1114-z RESEARCH ARTICLE Open Access In vitro application of ribonucleases: comparison of the effects on mRNA and miRNA stability Arian Aryani and Bernd Denecke* Abstract Background: MicroRNA has become important in a wide range of research interests. Due to the increasing number of known microRNAs, these molecules are likely to be increasingly seen as a new class of biomarkers. This is driven by the fact that microRNAs are relatively stable when circulating in the plasma. Despite extensive analysis of mechanisms involved in microRNA processing, relatively little is known about the in vitro decay of microRNAs under defined conditions or about the relative stabilities of mRNAs and microRNAs. Methods: In this in vitro study, equal amounts of total RNA of identical RNA pools were treated with different ribonucleases under defined conditions. Degradation of total RNA was assessed using microfluidic analysis mainly based on ribosomal RNA. To evaluate the influence of the specific RNases on the different classes of RNA (ribosomal RNA, mRNA, miRNA) ribosomal RNA as well as a pattern of specific mRNAs and miRNAs was quantified using RT-qPCR assays. By comparison to the untreated control sample the ribonuclease-specific degradation grade depending on the RNA class was determined. Results: In the present in vitro study we have investigated the stabilities of mRNA and microRNA with respect to the influence of ribonucleases used in laboratory practice. Total RNA was treated with specific ribonucleases and the decay of different kinds of RNA was analysed by RT-qPCR and miniaturized gel electrophoresis. -

Microrna Function: Multiple Mechanisms for a Tiny RNA?
Downloaded from rnajournal.cshlp.org on September 26, 2021 - Published by Cold Spring Harbor Laboratory Press REVIEW MicroRNA function: Multiple mechanisms for a tiny RNA? RAMESH S. PILLAI Friedrich Miescher Institute for Biomedical Research, 4002 Basel, Switzerland ABSTRACT MicroRNAs are sequence-specific regulators of post-transcriptional gene expression in many eukaryotes. They are believed to control the expression of thousands of target mRNAs, with each mRNA believed to be targeted by multiple microRNAs. Recent studies have uncovered various mechanisms by which microRNAs down-regulate their target mRNAs and have linked a well- known subcellular structure, the cytoplasmic processing bodies (PBs) to the microRNA pathway. The finding that microRNAs are misexpressed in cancers has reinforced the idea that their regulatory roles are very important. Keywords: microRNAs; miRNPs; translational repression; P-bodies; Argonaute INTRODUCTION that the forced overexpression of miRNAs can lead to the development of tumors (He et al. 2005). MicroRNAs (miRNAs) have come a long way from being Another pathway that uses small RNAs as sequence-spe- an oddity of worms when they were first discovered over 10 cific regulators is the RNA interference (RNAi) pathway, years ago (Lee et al. 1993) to be recognized now as novel which is an evolutionarily conserved response to the pres- agents exercising post-transcriptional control over most ence of double-stranded RNA (dsRNA) in the cell (Meister eukaryotic genomes. They are a family of 21–25-nucleotides and Tuschl 2004; Filipowicz 2005). The dsRNAs are cleaved (nt)-long RNAs expressed in a wide variety of organisms into 20-base pair (bp) duplexes of small-interfering RNAs ranging from plants to worms and humans. -

422.Full.Pdf
Downloaded from genome.cshlp.org on September 29, 2021 - Published by Cold Spring Harbor Laboratory Press Research Dioxin receptor and SLUG transcription factors regulate the insulator activity of B1 SINE retrotransposons via an RNA polymerase switch Angel Carlos Roma´n,1 Francisco J. Gonza´lez-Rico,1 Eduardo Molto´,2,3 Henar Hernando,4 Ana Neto,5 Cristina Vicente-Garcia,2,3 Esteban Ballestar,4 Jose´ L. Go´mez-Skarmeta,5 Jana Vavrova-Anderson,6 Robert J. White,6,7 Lluı´s Montoliu,2,3 and Pedro M. Ferna´ndez-Salguero1,8 1Departamento de Bioquı´mica y Biologı´a Molecular, Facultad de Ciencias, Universidad de Extremadura, 06071 Badajoz, Spain; 2Centro Nacional de Biotecnologı´a (CNB), Consejo Superior de Investigaciones Cientı´ficas (CSIC), Department of Molecular and Cellular Biology, Campus de Cantoblanco, C/Darwin 3, 28049 Madrid, Spain; 3Centro de Investigacio´n Biome´dica en Red de Enfermedades Raras (CIBERER), ISCIII, Madrid, Spain; 4Chromatin and Disease Group, Cancer Epigenetics and Biology Programme, Bellvitge Biomedical Research Institute (IDIBELL), Barcelona 08907, Spain; 5Centro Andaluz de Biologı´a del Desarrollo, CSIC-Universidad Pablo de Olavide, 41013 Sevilla, Spain; 6College of Medical, Veterinary and Life Sciences, University of Glasgow, Glasgow G12 8QQ, United Kingdom; 7Beatson Institute for Cancer Research, Glasgow, G61 1BD, United Kingdom Complex genomes utilize insulators and boundary elements to help define spatial and temporal gene expression patterns. We report that a genome-wide B1 SINE (Short Interspersed Nuclear Element) retrotransposon (B1-X35S) has potent in- trinsic insulator activity in cultured cells and live animals. This insulation is mediated by binding of the transcription factors dioxin receptor (AHR) and SLUG (SNAI2) to consensus elements present in the SINE. -

Transcriptomic Profiling of Equine and Viral Genes in Peripheral Blood
pathogens Article Transcriptomic Profiling of Equine and Viral Genes in Peripheral Blood Mononuclear Cells in Horses during Equine Herpesvirus 1 Infection Lila M. Zarski 1, Patty Sue D. Weber 2, Yao Lee 1 and Gisela Soboll Hussey 1,* 1 Department of Pathobiology and Diagnostic Investigation, Michigan State University, East Lansing, MI 48824, USA; [email protected] (L.M.Z.); [email protected] (Y.L.) 2 Department of Large Animal Clinical Sciences, Michigan State University, East Lansing, MI 48824, USA; [email protected] * Correspondence: [email protected] Abstract: Equine herpesvirus 1 (EHV-1) affects horses worldwide and causes respiratory dis- ease, abortions, and equine herpesvirus myeloencephalopathy (EHM). Following infection, a cell- associated viremia is established in the peripheral blood mononuclear cells (PBMCs). This viremia is essential for transport of EHV-1 to secondary infection sites where subsequent immunopathol- ogy results in diseases such as abortion or EHM. Because of the central role of PBMCs in EHV-1 pathogenesis, our goal was to establish a gene expression analysis of host and equine herpesvirus genes during EHV-1 viremia using RNA sequencing. When comparing transcriptomes of PBMCs during peak viremia to those prior to EHV-1 infection, we found 51 differentially expressed equine genes (48 upregulated and 3 downregulated). After gene ontology analysis, processes such as the interferon defense response, response to chemokines, the complement protein activation cascade, cell adhesion, and coagulation were overrepresented during viremia. Additionally, transcripts for EHV-1, EHV-2, and EHV-5 were identified in pre- and post-EHV-1-infection samples. Looking at Citation: Zarski, L.M.; Weber, P.S.D.; micro RNAs (miRNAs), 278 known equine miRNAs and 855 potentially novel equine miRNAs were Lee, Y.; Soboll Hussey, G. -

Exploring the Structure of Long Non-Coding Rnas, J
IMF YJMBI-63988; No. of pages: 15; 4C: 3, 4, 7, 8, 10 1 2 Rise of the RNA Machines: Exploring the Structure of 3 Long Non-Coding RNAs 4 Irina V. Novikova, Scott P. Hennelly, Chang-Shung Tung and Karissa Y. Sanbonmatsu Q15 6 Los Alamos National Laboratory, Los Alamos, NM 87545, USA 7 Correspondence to Karissa Y. Sanbonmatsu: [email protected] 8 http://dx.doi.org/10.1016/j.jmb.2013.02.030 9 Edited by A. Pyle 1011 12 Abstract 13 Novel, profound and unexpected roles of long non-coding RNAs (lncRNAs) are emerging in critical aspects of 14 gene regulation. Thousands of lncRNAs have been recently discovered in a wide range of mammalian 15 systems, related to development, epigenetics, cancer, brain function and hereditary disease. The structural 16 biology of these lncRNAs presents a brave new RNA world, which may contain a diverse zoo of new 17 architectures and mechanisms. While structural studies of lncRNAs are in their infancy, we describe existing 18 structural data for lncRNAs, as well as crystallographic studies of other RNA machines and their implications 19 for lncRNAs. We also discuss the importance of dynamics in RNA machine mechanism. Determining 20 commonalities between lncRNA systems will help elucidate the evolution and mechanistic role of lncRNAs in 21 disease, creating a structural framework necessary to pursue lncRNA-based therapeutics. 22 © 2013 Published by Elsevier Ltd. 24 23 25 Introduction rather than the exception in the case of eukaryotic 50 organisms. 51 26 RNA is primarily known as an intermediary in gene LncRNAs are defined by the following: (i) lack of 52 11 27 expression between DNA and proteins. -
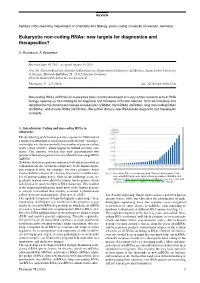
Eukaryotic Non-Coding Rnas: New Targets for Diagnostics and Therapeutics?
REVIEW Institute of Biochemistry, Department of Chemistry and Biology, Justus Liebig University of Giessen, Germany Eukaryotic non-coding RNAs: new targets for diagnostics and therapeutics? O. Rossbach, A. Bindereif Received June 30, 2015, accepted August 19, 2015 Prof. Dr. Albrecht Bindereif, Institute of Biochemistry, Department of Chemistry and Biology, Justus Liebig University of Giessen, Heinrich-Buff-Ring 58, 35392 Giessen, Germany [email protected] Pharmazie 71: 3–7 (2016) doi: 10.1691/ph.2016.5736 Non-coding RNAs (ncRNAs) in eukaryotes have recently developed to a very active research area in RNA biology, opening up new strategies for diagnosis and therapies of human disease. Here we introduce and describe the most important classes of eukaryotic ncRNAs: microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs). We further discuss new RNA-based diagnostic and therapeutic concepts. 1. Introduction: Coding and non-coding RNAs in eukaryotes The deciphering of the human genome sequence in 2000 marked a unique breakthrough in modern molecular biology. An impor- tant insight was the unexpectedly low number of protein-coding genes (about 20,000), which lagged far behind previous esti- mates. This number revealed that only approximately two percent of the human genome is transcribed into messenger RNA (mRNA). However, the human genome sequence itself did not provide an explanation for the enormous complexity of the human organ- ism compared with, for example, the more primitive worm Caenorhabditis elegans (C. elegans) that carries a similar num- Fig. 1: Non-coding RNAs as an emerging field. The rapid development of the ber of protein-coding genes. -

Autism Multiplex Family with 16P11.2P12.2 Microduplication Syndrome in Monozygotic Twins and Distal 16P11.2 Deletion in Their Brother
European Journal of Human Genetics (2012) 20, 540–546 & 2012 Macmillan Publishers Limited All rights reserved 1018-4813/12 www.nature.com/ejhg ARTICLE Autism multiplex family with 16p11.2p12.2 microduplication syndrome in monozygotic twins and distal 16p11.2 deletion in their brother Anne-Claude Tabet1,2,3,4, Marion Pilorge2,3,4, Richard Delorme5,6,Fre´de´rique Amsellem5,6, Jean-Marc Pinard7, Marion Leboyer6,8,9, Alain Verloes10, Brigitte Benzacken1,11,12 and Catalina Betancur*,2,3,4 The pericentromeric region of chromosome 16p is rich in segmental duplications that predispose to rearrangements through non-allelic homologous recombination. Several recurrent copy number variations have been described recently in chromosome 16p. 16p11.2 rearrangements (29.5–30.1 Mb) are associated with autism, intellectual disability (ID) and other neurodevelopmental disorders. Another recognizable but less common microdeletion syndrome in 16p11.2p12.2 (21.4 to 28.5–30.1 Mb) has been described in six individuals with ID, whereas apparently reciprocal duplications, studied by standard cytogenetic and fluorescence in situ hybridization techniques, have been reported in three patients with autism spectrum disorders. Here, we report a multiplex family with three boys affected with autism, including two monozygotic twins carrying a de novo 16p11.2p12.2 duplication of 8.95 Mb (21.28–30.23 Mb) characterized by single-nucleotide polymorphism array, encompassing both the 16p11.2 and 16p11.2p12.2 regions. The twins exhibited autism, severe ID, and dysmorphic features, including a triangular face, deep-set eyes, large and prominent nasal bridge, and tall, slender build. The eldest brother presented with autism, mild ID, early-onset obesity and normal craniofacial features, and carried a smaller, overlapping 16p11.2 microdeletion of 847 kb (28.40–29.25 Mb), inherited from his apparently healthy father. -

Activating Transcription Factor 6 Derepression Mediates Neuroprotection in Huntington Disease
Activating transcription factor 6 derepression mediates neuroprotection in Huntington disease José R. Naranjo, … , Jia-Yi Li, Britt Mellström J Clin Invest. 2016;126(2):627-638. https://doi.org/10.1172/JCI82670. Research Article Neuroscience Deregulated protein and Ca2+ homeostasis underlie synaptic dysfunction and neurodegeneration in Huntington disease (HD); however, the factors that disrupt homeostasis are not fully understood. Here, we determined that expression of downstream regulatory element antagonist modulator (DREAM), a multifunctional Ca2+-binding protein, is reduced in murine in vivo and in vitro HD models and in HD patients. DREAM downregulation was observed early after birth and was associated with endogenous neuroprotection. In the R6/2 mouse HD model, induced DREAM haplodeficiency or blockade of DREAM activity by chronic administration of the drug repaglinide delayed onset of motor dysfunction, reduced striatal atrophy, and prolonged life span. DREAM-related neuroprotection was linked to an interaction between DREAM and the unfolded protein response (UPR) sensor activating transcription factor 6 (ATF6). Repaglinide blocked this interaction and enhanced ATF6 processing and nuclear accumulation of transcriptionally active ATF6, improving prosurvival UPR function in striatal neurons. Together, our results identify a role for DREAM silencing in the activation of ATF6 signaling, which promotes early neuroprotection in HD. Find the latest version: https://jci.me/82670/pdf The Journal of Clinical Investigation RESEARCH ARTICLE Activating transcription factor 6 derepression mediates neuroprotection in Huntington disease José R. Naranjo,1,2 Hongyu Zhang,3 Diego Villar,1,2 Paz González,1,2 Xose M. Dopazo,1,2 Javier Morón-Oset,1,2 Elena Higueras,1,2 Juan C. -

A 1.37-Mb 12P11.22-P11.21 Deletion Coincident with a 367-Kb 22Q11.2
CORE Metadata, citation and similar papers at core.ac.uk Provided by Elsevier - Publisher Connector Taiwanese Journal of Obstetrics & Gynecology 53 (2014) 74e78 Contents lists available at ScienceDirect Taiwanese Journal of Obstetrics & Gynecology journal homepage: www.tjog-online.com Short Communication A 1.37-Mb 12p11.22ep11.21 deletion coincident with a 367-kb 22q11.2 duplication detected by array comparative genomic hybridization in an adolescent girl with autism and difficulty in self-care of menstruation Chih-Ping Chen a,b,c,d,e,f,*, Shuan-Pei Lin b,g,h,i, Schu-Rern Chern b, Peih-Shan Wu j, Jun-Wei Su a,k, Chen-Chi Lee a, Wayseen Wang b,l a Department of Obstetrics and Gynecology, Mackay Memorial Hospital, Taipei, Taiwan b Department of Medical Research, Mackay Memorial Hospital, Taipei, Taiwan c Department of Biotechnology, Asia University, Taichung, Taiwan d School of Chinese Medicine, College of Chinese Medicine, China Medical University, Taichung, Taiwan e Institute of Clinical and Community Health Nursing, National Yang-Ming University, Taipei, Taiwan f Department of Obstetrics and Gynecology, School of Medicine, National Yang-Ming University, Taipei, Taiwan g Department of Medicine, Mackay Medical College, New Taipei City, Taiwan h Department of Pediatrics, Mackay Memorial Hospital, Taipei, Taiwan i Mackay Junior College of Medicine, Nursing, and Management, Taipei, Taiwan j Gene Biodesign Co. Ltd, Taipei, Taiwan k Department of Obstetrics and Gynecology, China Medical University Hospital, Taichung, Taiwan l Department of Bioengineering, Tatung University, Taipei, Taiwan article info abstract Article history: Objective: To present an array comparative genomic hybridization (aCGH) characterization of a 12p11.22 Accepted 21 October 2013 ep11.21 microdeletion and 22q11.2 microduplication in an adolescent girl with autism, mental retar- dation, facial dysmorphism, microcephaly, behavior problems, and an apparently balanced reciprocal Keywords: translocation of t(8;12)(q24.3;p11.2). -

RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases
G C A T T A C G G C A T genes Review RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases Amber Willbanks, Shaun Wood and Jason X. Cheng * Department of Pathology, Hematopathology Section, University of Chicago, Chicago, IL 60637, USA; [email protected] (A.W.); [email protected] (S.W.) * Correspondence: [email protected] Abstract: Chromatin structure plays an essential role in eukaryotic gene expression and cell identity. Traditionally, DNA and histone modifications have been the focus of chromatin regulation; however, recent molecular and imaging studies have revealed an intimate connection between RNA epigenetics and chromatin structure. Accumulating evidence suggests that RNA serves as the interplay between chromatin and the transcription and splicing machineries within the cell. Additionally, epigenetic modifications of nascent RNAs fine-tune these interactions to regulate gene expression at the co- and post-transcriptional levels in normal cell development and human diseases. This review will provide an overview of recent advances in the emerging field of RNA epigenetics, specifically the role of RNA modifications and RNA modifying proteins in chromatin remodeling, transcription activation and RNA processing, as well as translational implications in human diseases. Keywords: 5’ cap (5’ cap); 7-methylguanosine (m7G); R-loops; N6-methyladenosine (m6A); RNA editing; A-to-I; C-to-U; 2’-O-methylation (Nm); 5-methylcytosine (m5C); NOL1/NOP2/sun domain Citation: Willbanks, A.; Wood, S.; (NSUN); MYC Cheng, J.X. RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases. Genes 2021, 12, 627. -

DNA Replication Stress Response Involving PLK1, CDC6, POLQ
DNA replication stress response involving PLK1, CDC6, POLQ, RAD51 and CLASPIN upregulation prognoses the outcome of early/mid-stage non-small cell lung cancer patients C. Allera-Moreau, I. Rouquette, B. Lepage, N. Oumouhou, M. Walschaerts, E. Leconte, V. Schilling, K. Gordien, L. Brouchet, Mb Delisle, et al. To cite this version: C. Allera-Moreau, I. Rouquette, B. Lepage, N. Oumouhou, M. Walschaerts, et al.. DNA replica- tion stress response involving PLK1, CDC6, POLQ, RAD51 and CLASPIN upregulation prognoses the outcome of early/mid-stage non-small cell lung cancer patients. Oncogenesis, Nature Publishing Group: Open Access Journals - Option C, 2012, 1, pp.e30. 10.1038/oncsis.2012.29. hal-00817701 HAL Id: hal-00817701 https://hal.archives-ouvertes.fr/hal-00817701 Submitted on 9 Jun 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution - NonCommercial - NoDerivatives| 4.0 International License Citation: Oncogenesis (2012) 1, e30; doi:10.1038/oncsis.2012.29 & 2012 Macmillan Publishers Limited All rights reserved 2157-9024/12 www.nature.com/oncsis ORIGINAL ARTICLE DNA replication stress response involving PLK1, CDC6, POLQ, RAD51 and CLASPIN upregulation prognoses the outcome of early/mid-stage non-small cell lung cancer patients C Allera-Moreau1,2,7, I Rouquette2,7, B Lepage3, N Oumouhou3, M Walschaerts4, E Leconte5, V Schilling1, K Gordien2, L Brouchet2, MB Delisle1,2, J Mazieres1,2, JS Hoffmann1, P Pasero6 and C Cazaux1 Lung cancer is the leading cause of cancer deaths worldwide. -

An HMG I/Y-Containing Repressor Complex and Supercolled DNA Topology Are Critical for Long-Range Enhancer-Dependent Transcription in Vitro
Downloaded from genesdev.cshlp.org on September 26, 2021 - Published by Cold Spring Harbor Laboratory Press An HMG I/Y-containing repressor complex and supercolled DNA topology are critical for long-range enhancer-dependent transcription in vitro Rajesh Bagga and Beverly M. Emerson 1 Regulatory Biology Laboratory, The Salk Institute for Biological Studies, La Jolla, California 92037 USA The 3' enhancer of the T cell receptor s.chain (TCR~) gene directs the tissue- and stage-specific expression and V(D)Jrecombination of this gene locus. Using an in vitro system that reproduces TCRoL enhancer activity efficiently, we show that long-range promoter-enhancer regulation requires a T cell-specific repressor complex and is sensitive to DNA topology. In this system, the enhancer functions to derepress the promoter on supercoiled, but not relaxed, templates. We find that the TCRoL promoter is inactivated by a repressor complex that contains the architectural protein HMG I/Y. In the absence of this repressor complex, expression of the TCR~ gene is completely independent of the 3' enhancer and DNA topology. The interaction of the T cell-restricted protein LEF-1 with the TCR~ enhancer is required for promoter derepression. In this system, the TCR~ enhancer increases the number of active promoters rather than the rate of transcription. Thus, long-range enhancers function in a distinct manner from promoters and provide the regulatory link between repressors, DNA topology, and gene activity. [Key Words: TCR genes; transcription; enhancers; HMG I/Y; derepression; DNA topology] Received December 27, 1996; revised version accepted January 14, 1997. The widespread importance of long-range promoter- Giaever 1988; Rippe et al.