Winter Wheat Adaptation to Climate Change in Turkey
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Roundup Ready Wheat – an Overview Based on Advancements in the Risk Assessment of Genetically Engineered Crops
Roundup Ready Wheat – An Overview Based on Advancements in the Risk Assessment of Genetically Engineered Crops by Doug Gurian-Sherman, Ph.D. Center for Science in the Public Interest 1875 Connecticut Avenue, N.W., Suite 300 Washington, DC 20009-5728 Phone: (202) 332-9110 www.cspinet.org TABLE OF CONTENTS SECTION PAGE Abstract................................................................................................................................. 2 Introduction.......................................................................................................................... 3 Background on the U.S. Regulatory System for GE Crops ............................................. 3 Characterization of the Transgene and Transgenic Protein............................................ 4 Human Safety....................................................................................................................... 6 Allergenicity ...................................................................................................................... 7 Unintended Adverse Effects.............................................................................................. 9 Environmental Issues ........................................................................................................ 11 Resistance Management ................................................................................................. 12 Gene Transfer.............................................................................................................. -

WINTER WHEAT Southern Idaho Dryland Winter Wheat Production Guide
SOUTHERN IDAHO DRYLAND WINTER WHEAT Southern Idaho Dryland Winter Wheat Production Guide Editors: Larry D. Robertson, Stephen O. Guy, and Bradford D. Brown BUL 827 1 SOUTHERN IDAHO DRYLAND WINTER WHEAT Southern Idaho Dryland Winter Wheat Production Guide Basic Recommendations • Winter wheat production can be improved and input costs reduced with good knowledge of growth and development. Learn to recognize the various growth stages and the impact of various management inputs. • Make an annual production management and marketing plan prior to beginning the crop season. • Minimize the number and intensity of tillage operations before and after winter wheat crops to control soil erosion, reduce water loss and soil compaction, and improve soil productivity. • Use rotations and cultural practices to minimize weed, disease, and insect problems, and reduce chemical use. • Choose varieties carefully with appropriate disease resistance, maturity, and quality characteristics for the intended use. • Prepare seedbeds carefully to conserve adequate moisture for germination and emergence, and to ensure good seed-soil contact. Seed at the proper time, depth, and rate for the chosen variety. • Use only high quality seed. Plant certified seed to ensure seed purity and viability. • Soil test to determine nutrient needs. Apply only the amounts of nutrients needed and at the proper time to avoid nutrient loss, wasted inputs, and environmental contamination. • Control weeds, insects, and diseases through variety choice, timely scouting, and application of the correct pesticides at the correct time and rate. • Plan ahead for storage and marketing needs. Become familiar with alternative marketing options. • Adjust combine properly to reduce kernel damage and dockage. • Store the crop in clean, insect-free bins, and check frequently for developing trouble spots. -

System for the Evaluation of Malting Quality of Wheat Varieties
KVASNÝ PRŮMYSL Kvasny prumysl (2020) 66: 232–238 https://doi.org/10.18832/kp2020.66.232 System for the evaluation of malting quality of wheat varieties Vratislav Psota*, Markéta Musilová Research Institute of Brewing and Malting Mostecká 971/7, 614 00 Brno Czech Republic *Corresponding author: [email protected] Abstract A new system for the calculation of malting quality of wheat varieties was designed. The malting parameters affected by a variety more than by a year or a site were used in the system. These parameters include wort viscosity, soluble nitrogen content, free amino nitrogen content, diastatic power, final attenuation of wort, and extract of malt. The sys- tem was used to assess the malting quality in 38 wheat varieties. The point value of each parameter was calculated using the regression equations, and the total value of the malting quality of wheat was determined by the modified “superiority measure” method. The results of the assessment of the wheat malting quality showed considerable dif- ferences between the varieties. Several varieties exhibited a more pronounced tendency to accumulate nitrogenous substances in grain. Some varieties, even with a slightly higher content of nitrogenous substances, provided above average malt quality. The set included the varieties that exhibited the optimal content of nitrogenous substances in grain, yet they did not give malt of good quality. Spring wheat varieties Alondra, Anabel, and Seance achieved signif- icantly above average malting quality. Dagmar, Evina, Sultan, Rumor, Sailor, and Brokat exhibited slightly above aver- age malting quality. The proposed system for monitoring malting quality of wheat varieties can help find the varieties suitable for the production of quality wheat malt. -

World Bank Document
Public Disclosure Authorized Key Agricultural Product Risk Assessment (KAPRA) Public Disclosure Authorized Public Disclosure Authorized Report on the Financial Resilience of Key Agricultural Products to Climate Change Public Disclosure Authorized 2018 We thank the Government of Japan and the World Bank Global Facility for Disaster Reduction and Recovery (GFDRR) for the valuable support they provided during the drafting and printing of this publication. This report has been drawn up by the Frankfurt School of Finance & Management for the World Bank. Responsibility for the content of this report rests solely with the Frankfurt School of Finance & Management, and does not necessarily reflect the opinions of the World Bank. Frankfurt School of Finance & Management gGmbH Adickesallee 32–34, 60322 Frankfurt am Main [email protected] http://www.frankfurt-school.de Table of Contents Tables .......................................................................................................................................................................................... 4 Figures ........................................................................................................................................................................................ 4 Annexes ...................................................................................................................................................................................... 5 Abbreviations ............................................................................................................................................................................. -
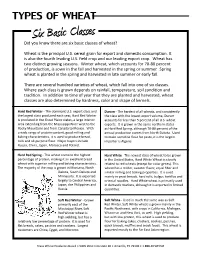
TYPES of WHEAT Six Basic Classes Did You Know There Are Six Basic Classes of Wheat?
TYPES OF WHEAT Six Basic Classes Did you know there are six basic classes of wheat? Wheat is the principal U.S. cereal grain for export and domesti c consumpti on. It is also the fourth leading U.S. fi eld crop and our leading export crop. Wheat has two disti nct growing seasons. Winter wheat, which accounts for 70-80 percent of producti on, is sown in the fall and harvested in the spring or summer. Spring wheat is planted in the spring and harvested in late summer or early fall. There are several hundred varieti es of wheat, which fall into one of six classes. Where each class is grown depends on rainfall, temperature, soil conditi on and traditi on. In additi on to ti me of year that they are planted and harvested, wheat classes are also determined by hardness, color and shape of kernels. Hard Red Winter - The dominant U.S. export class and Durum - The hardest of all wheats, and consistently the largest class produced each year, Hard Red Winter the class with the lowest export volume, Durum is produced in the Great Plains states, a large interior accounts for less than 5 percent of all U.S. wheat area extending from the Mississippi River west to the exports. It is grown in the same northern states Rocky Mountains and from Canada to Mexico. With as Hard Red Spring, although 70-80 percent of the a wide range of protein content, good milling and annual producti on comes from North Dakota. Used baking characteristi cs, it is used to produce bread, to make semolina fl our for pasta, it is the largest rolls and all-purpose fl our. -

International Grains Council
www.igc.int IGCIGC 20112013 igc.int International Grains Council AMIS Crops: Market Situation and Outlook in 2013/14 Third Session of the AMIS Global Food Market Information Group Paris, OECD Headquarters 23-24 April 2013 Amy Reynolds International Grains Council [email protected] Exporter stocks tighten in 2012/13, www.igc.int IGCIGC 20112013 good 2013 crops needed to replenish supplies igc.int Maize: Soyabeans: Wheat: m t m t Black Sea leads decline m t US lowest since 95/96 Total up, but US still down 90 70 25 Others Argentina Argentina EU Ukraine Brazil 80 Black Sea 60 Brazil US US US 20 70 50 60 15 40 50 40 30 10 30 20 20 5 10 10 0 0 0 03/04 04/05 05/06 06/07 07/08 08/09 09/10 10/11 11/12 12/13 03/04 04/05 05/06 06/07 07/08 08/09 09/10 10/11 11/12 12/13 03/04 04/05 05/06 06/07 07/08 08/09 09/10 10/11 11/12 12/13 1 www.igc.int First fall in total grains consumption in 14 years; IGCIGC 20112013 igc.int lower wheat feeding and maize industrial use m t y/y change 20% Food Food Industrial 2000 Feed Feed Other 15% Industrial 1500 10% 1000 5% 500 0% 0 -5% www.igc.int IGCIGC 20112013 IGC GOI down from highs, but grains still elevated igc.int IGC daily index (GOI), rebased: Jan 2012 = 100 Dow Jones–UBS commodity sub-indices 160 150 Y/Y Grains Wheat +10.4% 150 140 Prec. -

This Crisp and Refreshing Cider Mixes the Sweetness of the Apples with a Subtle Dryness for a Balanced Cider Taste. a Refreshing
o Live Team Trivia at 8PM just Tap’d: Harpoon ‘Leviathan’ & Kentucky ‘Peach Barrel’ Try the Firecracker Shrimp or Angus Sliders w/ Tap Fries! This crisp and refreshing cider mixes the sweetness of the apples with a subtle dryness for a balanced cider taste. A refreshing mix of malted wheat flavor and fruity notes, wrapped up in a distinctively citrusy hop aroma. Brewed around the concept of balance. Bursting with hop aromas ranging from pine to grapefruit notes. Two Hearted remains one of the defining American style India Pale Ales. Pleasant fruity, floral hop aroma & earthy flavor perfectly balance the mild sweetness of the English grains. Brewed with the nectar of the great Salmiana Agave. Agave complements the refreshingly light quality of this Wheat Ale and adds a subtle note of flavor. Begins with a complex biscuit & dark toast aroma. Seven different malts with balancing hops round out the medium dry finish. An all malt wheat ale infused with real lemons and limes. This shandy has a lively fruity flavor and a powerful "throw your head back" fresh lemon aroma. A bounty of American hops delivers the signature clean and crisp finish of a perfectly balanced West Coast style IPA. An abundance of American hops balanced with a hint of malt to produce an exhilarating experience of intense hop flavors and aromas. A session IPA naturally brewed with a complex array of malts, grains and hops. Balanced for optimal aromatic and a clean finish. Hop bitterness is nicely balanced with the malt. Highlighted with soft notes of chocolate and slight undertones of roasted coffee. -

2017 Regional Quality Survey
Hard Red Winter Wheat 2017 Regional Quality Survey PHOTO CREDIT: KIMBERLY WARNER 127 Noble Research Center Stillwater, OK 74078 ph 405.744.9333 [email protected] www.plainsgrains.org Idaho Wheat Commission Colorado Wheat Oklahoma Wheat Commission www.idahowheat.org Administrative Committee www.wheat.state.ok.us www.coloradowheat.org North Dakota South Dakota Kansas Wheat Commission Wheat Commission Wheat Commission www.kswheat.com www.ndwheat.com www.sdwheat.org Washington Grain Commission Texas Wheat Producers Nebraska Wheat Board www.washingtongrainalliance.com Board and Association www.nebraskawheat.com www.texaswheat.org Montana Wheat & Barley Wyoming Wheat Committee Oregon Wheat Commission Growers Association wbc.agr.mt.gov www.owgl.org www.wyomingwheat.com 2017 Hard Red Winter Wheat Regional Quality Survey Plains Grains, Inc. Plains Grains, Inc., a non-profit, private marketing goals to have quality data for quality based marketing initiative, was the entire HRWP wheat production G area. I. formed in 2004 through the OklahomaPGI Each state may be able to produce the Wheat Commission, the Oklahoma quality needed by foreign buyers, but it Department of Agriculture, Food will take multiple states to achieve the and Forestry, the Oklahoma critical mass needed to meet the State University Division quantity needs. By working of Agricultural Sciences together as a region we and Natural Resources. can meet both quality and quantity demands. PGI was designed to bridge the gap PGI In 2004, PGI’s crop between wheat quality survey included producers, grain P G I. the Oklahoma HRW companies and foreign wheat crop. Designed as and domestic flour millers a regional marketing entity, to benefit all segments of the wheat PGI then brought five other HRW wheat industry. -

High-Protein Nutritious Flatbreads and an Option for Gluten-Sensitive Individuals
foods Article High-Protein Nutritious Flatbreads and an Option for Gluten-Sensitive Individuals Talwinder S. Kahlon *, Roberto J. Avena-Bustillos, Jenny L. Brichta and Ashwinder K. Kahlon Western Regional Research Center, USDA-ARS, 800 Buchanan St., Albany, CA 94710, USA; [email protected] (R.J.A.-B.); [email protected] (J.L.B.); [email protected] (A.K.K.) * Correspondence: [email protected]; Tel.: +1-510-559-5665 or +1-510-559-5660 Received: 26 September 2019; Accepted: 15 November 2019; Published: 19 November 2019 Abstract: Whole grain quinoa and wheat, high-protein vegetable flatbreads were evaluated by tasters and a physical analysis was conducted. The objective was to produce nutritious, tasty gluten-free (quinoa) as well as gluten-containing (wheat) flatbreads. Flatbreads were Quinoa Peanut Oilcake Broccoli (QPCBROC), Wheat Peanut Oilcake Broccoli (WPCBROC), Quinoa Peanut Oilcake Beets (QPCBEET) and Wheat Peanut Oilcake Beets (WPCBEET). Peanut Oilcake would increase protein and add value to this farm byproduct. Bile acid binding broccoli and beets with cholesterol-lowering potential were used. Tasters preferred QPCBROC flatbreads for all sensory parameters. Acceptance of flatbreads was QPCBROC (83%), WPCBROC (70%), QPCBEET (78%) and WPCBEET (69%); these values were statistically similar. The objective of 25% protein content was exceeded by 5–8% and that ≥ of 70% acceptance was adequately achieved. These flatbreads were low in fat (5–6%) and contained ≥ essential minerals (4%) with only 1% added salt. Porosity and expansion data suggest that these ≤ flatbreads would take up relatively little shelf space. These flatbreads require only three ingredients and can be made in a household kitchen or by commercial production. -

Crop Profile for Hard Red Spring and Durum Wheats in North Dakota
Crop Profile for Hard Red Spring and Durum Wheats in North Dakota Prepared June, 2000 General Production Information The primary production region for durum is the Northwest and North Central part of the state. Of 3,000,000 acres of durum planted in 1998, 1,520,000 were planted in the Northwest district, 410,000 in the West Central, and 370,000 in the North Central. Spring wheat was grown statewide. Of the 6,700,000 acres of spring wheat planted in 1998, 1,250,000 were planted in the Northeast district, 990,000 in the East Central, and 830,000 in the Southeast. Table 1. Wheat Production and its Economic impact on North Dakotas Economy National Total Acres Total Production Cash Value Year Crop Rank Planted (bushels) ($) Durum 1st 3,000,000 79,380,000 447,120,000 1996 Spring Wheat 1st 9,600,000 313,500,000 1,269,675,000 Durum 1st 2,700,000 56,540,000 284,093,000 1997 Spring wheat 1st 8,800,000 210,000,000 730,800,000 Durum 1st 3,000,000 97,350,000 311,520,000 1998 Spring wheat 1st 6,700,000 211,200,000 665,280,000 Durum 1st 3,450,000 72,000,000 --- 1999 Spring Wheat 1st 5,900,000 168,000,000 --- Cultural Practices North Dakota has a temperate climate that is conducive to growing wheat. Average annual days above freezing range from 110 days in the North to 130 days in the South. Average growing season precipitation ranges from 16.0 inches in the Southeast to less than 12.0 inches in the Northwest. -

The Analysis of Climate Change Awareness at Local Level İklim
Avrupa Bilim ve Teknoloji Dergisi European Journal of Science and Technology Sayı 25, S. 457-462, Ağustos 2021 No. 25, pp. 457-462, August 2021 © Telif hakkı EJOSAT’a aittir Copyright © 2021 EJOSAT Araştırma Makalesi www.ejosat.com ISSN:2148-2683 Research Article The Analysis of Climate Change Awareness at Local Level Burçin Atılgan Türkmen1* 1* Bilecik Seyh Edebali University, Faculty of Engineering, Departmant of Chemical Engineering, Bilecik, Turkey, (ORCID: 0000-0003-3220-3817), [email protected] (First received 4 March 2021 and in final form 26 June 2021) (DOI: 10.31590/ejosat.890865) ATIF/REFERENCE: Atilgan Turkmen, B., (2021). The Analysis of Climate Change Awareness at Local Level. European Journal of Science and Technology, (25), 457-462. Abstract Climate change is one of the biggest problems facing the world today due to its negative effects on human health and ecosystems. It is important to evaluate society's awareness of climate change to avoid this problem. Understanding the drivers of climate change attitudes is critical, as an action to address climate change is dependent on public awareness. Therefore, this research was carried out to identify the level of awareness and knowledge of climate change of residents in the province of Bilecik. In order to collect data, a questionnaire consisting of 11 questions was applied to randomly selected 508 people. Statistical approaches have been used to test the data collected from questionnaires. According to the results of the research, of the respondents 96% stated that they believe that climate change is occurring, 89% stated that climate change is occurring in Bilecik, of the respondents 87% believe that climate change is threatening. -

Subpart M -- United States Standards for Wheat
United States Department of Agriculture Grain Inspection, Packers and Stockyards Administration Federal Grain Inspection Service U.S. Standards May 2014 Subpart M -- United States Standards for Wheat Contents Subpart M -- United States Standards for Wheat 1 Terms Defined 2 Principles Governing the Application of Standards 4 Grades and Grade Requirements 5 Special Grades and Special Grade Requirements 6 The U.S. Department of Agriculture (USDA) prohibits discrimination in its programs on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, and marital or familial status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternate means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD). To file a complaint, write to the USDA, Office of Civil Rights, Room 326-W, 1400 Independence Avenue, SW, Washington, DC 20250-9410, or call (202) 720-5964 (voice and TDD). USDA is an equal employment opportunity employer. Distribution: FGIS, OSP, Industry Originating Office: FMD PPMAB Terms Defined § 810.2201 Definition of wheat. Grain that, before the removal of dockage, consists of 50 percent or more common wheat (Triticum aestivum L.), club wheat (T. compactum Host.), and durum wheat (T. durum Desf.) and not more than 10 percent of other grains for which standards have been established under the United States Grain Standards Act and that, after the removal of the dockage, contains 50 percent or more of whole kernels of one or more of these wheats. § 810.2202 Definition of other terms.