Number 38 December 2012
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Development of Forest Sector in the Arkhangelsk Oblast During the Transition Period of the 1990S
Development of forest sector in the Arkhangelsk oblast during the transition period of the 1990s ALBINA PASHKEVICH Pashkevich Albina (2003). Development of forest sector in the Arkhangelsk oblast during the transition period of the 1990s. Fennia 181: 1, pp. 13–24. Helsinki. ISSN 0015-0010. The Arkhangelsk oblast has long been one of Russia’s most important forest industrial regions. This paper analyses the changes in accessibility of forest resources and forest commodity production during the transition period in the 1990s. Special attention is given to firm restructuring, active roles of domestic capital and the different survival strategies that have been developed by in- dustries in the region. Further analysis deals with signs of economic recovery in the forest sector due to the processes of restructuring, modernisation and self-organisation. Albina Pashkevich, Spatial Modelling Centre (SMC), Department of Social and Economic Geography, Umeå University, Box 839, SE-98128 Kiruna, Sweden. E-mail: [email protected]. MS received 12 August 2002. Introduction adoption of a new. Some suggest that this proc- ess has been deeply embedded in the nature of The shift from central planning to a market-based the socialist system (Dingsdale 1999; Hamilton economy in Russia culminated with the dramatic 1999) and that the legacy of the communism has economic and political reorientation that began been only partly removed, and instead has mere- in the 1990s. This transition towards a market-ori- ly been reworked in a complex way (Smith 1997). ented and outward-looking economic system led Others say that reforms have actually ended the by private sector has created new challenges and old ‘command economy’ but have instead suc- opportunities. -

RCN #33 21/8/03 13:57 Page 1
RCN #33 21/8/03 13:57 Page 1 No. 33 Summer 2003 Special issue: The Transformation of Protected Areas in Russia A Ten-Year Review PROMOTING BIODIVERSITY CONSERVATION IN RUSSIA AND THROUGHOUT NORTHERN EURASIA RCN #33 21/8/03 13:57 Page 2 CONTENTS CONTENTS Voice from the Wild (Letter from the Editors)......................................1 Ten Years of Teaching and Learning in Bolshaya Kokshaga Zapovednik ...............................................................24 BY WAY OF AN INTRODUCTION The Formation of Regional Associations A Brief History of Modern Russian Nature Reserves..........................2 of Protected Areas........................................................................................................27 A Glossary of Russian Protected Areas...........................................................3 The Growth of Regional Nature Protection: A Case Study from the Orlovskaya Oblast ..............................................29 THE PAST TEN YEARS: Making Friends beyond Boundaries.............................................................30 TRENDS AND CASE STUDIES A Spotlight on Kerzhensky Zapovednik...................................................32 Geographic Development ........................................................................................5 Ecotourism in Protected Areas: Problems and Possibilities......34 Legal Developments in Nature Protection.................................................7 A LOOK TO THE FUTURE Financing Zapovedniks ...........................................................................................10 -

2018 FIFA WORLD CUP RUSSIA'n' WATERWAYS
- The 2018 FIFA World Cup will be the 21st FIFA World Cup, a quadrennial international football tournament contested by the men's national teams of the member associations of FIFA. It is scheduled to take place in Russia from 14 June to 15 July 2018,[2] 2018 FIFA WORLD CUP RUSSIA’n’WATERWAYS after the country was awarded the hosting rights on 2 December 2010. This will be the rst World Cup held in Europe since 2006; all but one of the stadium venues are in European Russia, west of the Ural Mountains to keep travel time manageable. - The nal tournament will involve 32 national teams, which include 31 teams determined through qualifying competitions and Routes from the Five Seas 14 June - 15 July 2018 the automatically quali ed host team. A total of 64 matches will be played in 12 venues located in 11 cities. The nal will take place on 15 July in Moscow at the Luzhniki Stadium. - The general visa policy of Russia will not apply to the World Cup participants and fans, who will be able to visit Russia without a visa right before and during the competition regardless of their citizenship [https://en.wikipedia.org/wiki/2018_FIFA_World_Cup]. IDWWS SECTION: Rybinsk – Moscow (433 km) Barents Sea WATERWAYS: Volga River, Rybinskoye, Ughlichskoye, Ivan’kovskoye Reservoirs, Moscow Electronic Navigation Charts for Russian Inland Waterways (RIWW) Canal, Ikshinskoye, Pestovskoye, Klyaz’minskoye Reservoirs, Moskva River 600 MOSCOW Luzhniki Arena Stadium (81.000), Spartak Arena Stadium (45.000) White Sea Finland Belomorsk [White Sea] Belomorsk – Petrozavodsk (402 km) Historic towns: Rybinsk, Ughlich, Kimry, Dubna, Dmitrov Baltic Sea Lock 13,2 White Sea – Baltic Canal, Onega Lake Small rivers: Medveditsa, Dubna, Yukhot’, Nerl’, Kimrka, 3 Helsinki 8 4,0 Shosha, Mologa, Sutka 400 402 Arkhangel’sk Towns: Seghezha, Medvezh’yegorsk, Povenets Lock 12,2 Vyborg Lakes: Vygozero, Segozero, Volozero (>60.000 lakes) 4 19 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 30 1 2 3 6 7 10 14 15 4,0 MOSCOW, Group stage 1/8 1/4 1/2 3 1 Estonia Petrozavodsk IDWWS SECTION: [Baltic Sea] St. -

Материалы Совещания Рабочей Группы INQUA Peribaltic
САНКТ-ПЕТЕРБУРГСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ Материалы совещания рабочей группы INQUA Peribaltic Из сборника материалов совместной международной конференции «ГЕОМОРФОЛОГИЯ И ПАЛЕОГЕОГРАФИЯ ПОЛЯРНЫХ РЕГИОНОВ», симпозиума «Леопольдина» и совещания рабочей группы INQUA Peribaltic, Санкт-Петербург, СПбГУ, 9 – 17 сентября 2012 года Санкт-Петербург, 2012 SAINT-PETERSBURG STATE UNIVERSITY Proceedings of the INQUA Peribaltic Working Group Workshop From the book of proceeding of the Joint International Conference “GEOMORPHOLOGY AND PALАEOGEOGRAPHY OF POLAR REGIONS”, Leopoldina Symposium and INQUA Peribaltic Working Group Workshop, Saint-Petersburg, SPbSU, 9-17 September, 2012 Saint-Petersburg, 2012 УДК 551.4 Ответственные редакторы: А.И. Жиров, В.Ю. Кузнецов, Д.А. Субетто, Й. Тиде Техническое редактирование и компьютерная верстка: А.А. Старикова, В.В. Ситало Обложка: К.А. Смыкова «ГЕОМОРФОЛОГИЯ И ПАЛЕОГЕОГРАФИЯ ПОЛЯРНЫХ РЕГИОНОВ»: Материалы совместной международной конференции «ГЕОМОРФОЛОГИЯ И ПАЛЕОГЕОГРАФИЯ ПОЛЯРНЫХ РЕГИОНОВ», симпозиума «Леопольдина» и совещания рабочей группы INQUA Peribaltic. Санкт-Петербург, СПбГУ, 9 – 17 сентября 2012 года / Отв. ред. А.И. Жиров, В.Ю. Кузнецов, Д.А. Субетто, Й. Тиде. – СПб., 2012. – 475 с. ISBN 978-5-4391-0029-3 Сборник содержит материалы совместной международной конференции "Геоморфологические и палеогеографические исследования полярных регионов", симпозиума «Леопольдина» и совещания рабочей группы INQUA Peribaltic. Обсуждается целый ряд актуальных вопросов, связанных с изучением проблем теоретической -

Transport and Infrastructural Basis of the Tourism Development Strategy in the Arkhangelsk Oblast © Aleksandr Yu
Aleksandr Yu. TSVETKOV. Transport and infrastructural basis … 35 UDC [338.48+332.14](470.11)(045) DOI: 10.37482/issn2221-2698.2020.38.44 Transport and infrastructural basis of the tourism development strategy in the Arkhangelsk Oblast © Aleksandr Yu. TSVETKOV, Cand. Sci. (Econ.), associate professor E-mail: [email protected] Department of Management, Higher School of Economics, Management and Law, Northern (Arctic) Federal University named after M.V. Lomonosov, Arkhangelsk, Russia Abstract. The article, devoted to the analysis of transport and geographical locations, describes possible strategies for the development of tourism in the Arkhangelsk Oblast. The main goal of the research was the development of logistic schemes of the transportation of tourists from the places of formation of tourist flows to the Arkhangelsk Oblast. The methodological basis of the article is to determine the economic dis- tances between potential tourist distribution centers and their places of interest in the area. Moscow, St. Petersburg, and Arkhangelsk were considered as the main towns of departure. Kargopol, Solvychegodsk, Kholmogory and Lomonosovo, Solovki, Kenozersky National Park, and Pinega caves are regarded as the main sites of tourist interest in the Arkhangelsk Oblast. It was determined that Kargopol is the most acces- sible for tourists, and Kenozersky National Park is the most recognizable by tourists but the least accessible. The object of world cultural heritage, the Monastery of the Transfiguration of the Saviour on Solovki is the most accessible for tourists from the territory of Karelia. It is recommended to optimize the schedule and to synchronize the work of transport for tourists to improve the transport accessibility of recreational facili- ties in the area. -

Assessment of Organic Matter Temporal Dynamics in the Klyazma Basin Using Remote Sensing and Qgis Trends.Earth
ISSN-E 1995-9516 Universidad Nacional de Ingeniería COPYRIGHT © (UNI). TODOS LOS DERECHOS RESERVADOS http://revistas.uni.edu.ni/index.php/Nexo https://doi.org/10.5377/nexo.v34i02.11624 Vol. 34, No. 02, pp. 973-992/Junio 2021 Assessment of organic matter temporal dynamics in the klyazma basin using remote sensing and qgis trends.earth Evaluación de la dinámica temporal de la materia orgánica en la cuenca de klyazma utilizando monitoreo remoto y qgis trends.earth Tatiana A. Trifonova1, Natalia V. Mishchenko2,*, Pavel S. Shutov2 1 Department of Soil Science, Lomonosov Moscow State University, Moscow, Russian Federation 2 Institute of Biology and Ecology, Vladimir State University, Vladimir, Russian Federation * [email protected] (recibido/received: 15-febrero-2021; aceptado/accepted: 05-abril-2021) ABSTRACT The article addresses the dynamics of biological processes in various landscapes within a holistic natural geosystem—a catchment area. The Klyazma river (the fourth order tributary to the Volga) was selected as the object of study. The natural complex of the Klyazma river basin is a combination of different landscapes, each marked by a diverse composition of geomorphological and soil-vegetation structures. The study is based on remote sensing data and the Trends.Earth Land Degradation Monitoring Project (Land Cover Dataset, European Space Agency 2015, 300 m spatial resolution) implemented using the open-source Quantum GIS 2.18. Four landscape provinces and eight site were identified in the studied catchment area according to the geomorphological structure and the soil and vegetation cover. The ecosystem parameters Gross Primary Productivity, Net Primary Productivity, and Ecosystem Respiration were measured in the identified sites. -

Download Article (PDF)
Advances in Engineering Research, volume 177 International Symposium on Engineering and Earth Sciences (ISEES 2018) Basin Approach in Geomorphological Studies of Klyazma River Basin Structure Roman Vladimirovich Repkin Alla Valeryevna Lyubesheva Vladimir State University, Vladimir State University, Department of Biology and Ecology, Department of Biology and Ecology, Vladimir, Vladimir Region, Russia Vladimir, Vladimir Region, Russia [email protected] [email protected] Ekaterina Lvovna Pronina Vladimir State University, Department of Biology and Ecology, Vladimir, Vladimir Region, Russia [email protected] Abstract – This work is devoted to geomorphological studies of Т.А. also developed a basin approach for the functioning of the structures of the Klyazma River basin and factors of modern small river basins for mountain areas and for lowland rivers [7, exogenous morphogenesis in the river basin and its morphological 9]. A wide and detailed description of the Klyazma River basin parts. The landscapes of the basins of small rivers have been and its tributaries is being conducted at the Department of experiencing anthropogenic load for a long time, so the processes Biology and Ecology of the VlSU under the direction of Dr. of relief formation have their own specifics. To create a geomorphological model of the transformation of landscapes of T.A. Trifonova. the Klyazma River river basin with the aim of ecological forecasting and monitoring, the data were obtained on the II. METHODS AND MATERIALS dynamics of exogenous processes, among which the most common are: karst, hydromorphism, gully and lateral erosion, landslide With the use of remote, statistical, mathematical methods formation. and GIS technologies, the selection of sites for the imposition of geomorphological profiles was carried out on the basis of Keywords – component; formatting; style; styling; insert analysis of space images of different resolution and analysis of the conjugated data of the studies. -

Download Article (PDF)
Advances in Social Science, Education and Humanities Research, volume 144 3rd International Conference on Arts, Design and Contemporary Education (ICADCE 2017) Rituals and Customs as a Reflection of the Folk Tradition The Russian North in the Context of Culture Magda Djichonaya Institute of Slavic Culture Russian State University named after A.N. Kosygin Moscow, Russia E-mail: [email protected] Abstract—The article is devoted to folklore traditions and is II. TRADITIONAL HOUSEHOLD WAY OF LIFE OF considered by the example of the Russian North. Ethno- INHABITANTS OF THE ARKHANGELSK REGION cultural realities reflect the specifics of the structure of artistic culture and of the existence in it of authentic folklore. Russia‘s Arkhangelsk Region, for instance, due to its remoteness, is distinguished by preserved in the area unique Keywords—polyfunctionality of art; art in the context of centuries-old folk traditions. Its art presents the masterpiece culture; folk traditions; ethnic and cultural realities of people and the perception in the mass. Folklore traditions are very well preserved, as handed down from generation to I. INTRODUCTION generation. The specificity of the cultural and logical approach to the Description of nature, navigation, hunting, fishing, study of art lies in its consideration as part of a holistic socio- peculiarities of female labor, buildings, clothing, etc. cultural space as a special sphere of human activity and, contained numerous realities of everyday life of the fishing finally, as a cultural phenomenon in which the artistic and population of the Russian North, from which came the aesthetic values are accumulated. storytellers. Openness to artistic culture as a system lies in its ability The key elements of the way of life of northerners — to restrict and expand one‘s frame of graduation. -
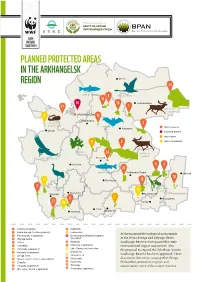
Planned Protected Areas in the Arkhangelsk Region
PLANNED PROTECTED AREAS IN THE ARKHANGELSK Mezen REGION 23 atlantic salmon 8 whooper swan 13 16 14 Leshukonskoe 17 1 wild forest reindeer ARKHANGELSK 9 Новодвинск 12 Kholmogory freshwater 3 pearl mussel вrandt's nature reserve bat Karpogory Onega botanical garden nature park 11 2 nature monument tree lungwort 24 wild forest reindeer white-tailed eagle Bereznik Plesetsk 4 4 22 atlantic salmon Verkhnyaya Toyma Yarensk 10 15 Shenkursk 18 5 common lady’s slipper crane 19 orchid Nyandoma Kargopol Krasnoborsk 7 6 Koryazhma 21 20 Котлас Velsk Konosha Oktyabrsky 1 Verkhnechelassky 14 Sebboloto 2 15 Dvina-Pinega (Verkhneyulovsky) Lekshmokh At the moment the ecological assessments 3 Puchkomsky (expansion) 16 Dendrological (botanical) garden 4 Uftyuga-Ilesha “SevNIILH” of the Dvina-Pinega and Uftyuga-Ilesha 5 Atleka 17 Solzinsky Landscape Reserves have passed the state 6 Voloshsky 18 Shilovsky (expansion) environmental impact assessment. Also 7 Ustiansky (expansion) 19 Lake Churozero (correction the proposal to expand the Zelezhnye Vorota 8 Kuloysky (expansion) of borders) 20 Landscape Reserve has been approved. These 9 Onega coast Turovsky Les 21 10 Nature reserve in the Lensky district Kovzhinsky documents determine, among other things, 22 11 Zvozsky Soyginsky the borders, protection regimes and 23 12 Chugsky (expansion) Timansky conservation zones of the nature reserves. 24 13 Zheleznye Vorota (expansion) Klonovsky (expansion) Intact forest landscapes are Arkhangelsk Region has been shaped by optimal habitats for such coniferous forests, covering most of its animals as bears, pine martens, elks and territory. Its virgin forests, which have not capercaillies. been disturbed by human activities, play a key role in protecting populations of rare and threatened species. -

Mikhail Zhilin EARLY MESOLITHIC BONE ARROWHEADS from the VOLGA-OKA INTERFLUVE, CENTRAL RUSSIA
Fennoscandia archaeologica XXXII (2015) Mikhail Zhilin EARLY MESOLITHIC BONE ARROWHEADS FROM THE VOLGA-OKA INTERFLUVE, CENTRAL RUSSIA Abstract Several different types of bone arrowheads were produced and used during the Early Mesolithic in the Volga-Oka interfluve. In this paper, recent research on these artefacts is reviewed, and their means of manufacture and ways of use determined through microscopy and experimental research. The research highlights the skill of the Early Mesolithic inhabitants of the Volga-Oka interfluve in manufacturing bone arrowheads, used for hunting various animals. A similarity was observed between the flint industry of pre-boreal sites of the Volga-Oka region and those of southern Finland, especially with respect to tanged flint arrowheads. The similarity suggests that the Early Mesolithic population of southern Finland and Karelian Isthmus most probably produced and used bone arrowheads similar to those described in the article. However, because bone artefacts are generally missing in the latter regions because of acrid soils, the conclusion should be regarded as a working hypothesis. Keywords: Early Mesolithic, bone, projectile points, Upper Volga, eastern Baltic, southern Finland Mikhail Zhilin, Department of Stone Age Archaeology, Institute of Archaeology, Russian Academy of Sciences, ul. Planernaya 3-2-235, RU-125480 Moscow, Russia: [email protected]. Received: 24 May 2015; Accepted: 29 Aug 2015; Revised: 5 Sep 2015 INTRODUCTION to c 9500–9200 BP have produced artefacts made of local lithic raw materials, mainly quartz, as Excavations of peat bog sites in the Volga-Oka well as imported high-quality Cretaceous and interfluve during the last three decades have Carboniferous fl int (Takala 2004; Hertell & Tal- produced rich lithic, bone and antler industry lavaara 2011; Manninen & Hertell 2011). -

Tver State Medical Academy,Russia
Welcome to Tver State Medical Academy Bazhenov D.V. Vice Chancellor for international students Welcome to our medical academy, an institution replete with tradition, values, enthusiasm, quality and international prestige. Our educational model is student-oriented. Opportunities to learn for yourself and to participate in a research project are highly recommended. Throughout your medical training you will witness many very rich opportunities to exchange opinions and ideas with classmates from different countries, thus increasing your awareness of cultural diversity and placing you on the correct tract to treat patients of different cultures. In our renowned Program of Medicine in the Community, opportunities to increase your clinical skills abound. Your participation in supporting the underserved population will be enormously appreciated and of immense benefit to you. You will be one of the foreign students who develop, practice and master bilingual skills while also developing skills related to humanism, professionalism and altruism. The Tver State Medical Academy works arduously to assure that at the end of your training you will be a physician with a special profile: a knowledgeable, dutiful, clinician, communicator, researcher, and long-term learner for the benefit of your community. he City Tver Central Federal District Tver Region Tver region is the largest of the 11 regions of the central Russia. Its territory is 84100 square kms and it includes 36 administrative districts, 23 towns & 8941 small villages. Population of Tver region is 1.6 million people. The City of Tver ( formerly called Kalinin from 1931 to 1990 ) was founded in 1246, it was the capital of a powerful medieval state and later the typical provincial town of the Russian Empire. -

Demographic Potential of the Klyazma River Basin-History of Development and Present State
Modern Applied Science; Vol. 9, No. 2; 2015 ISSN 1913-1844 E-ISSN 1913-1852 Published by Canadian Center of Science and Education Demographic Potential of the Klyazma River Basin-History of Development and Present State Trifonova Tatiana Anatolievna1 & Repkin Roman Vladimirovich2 1 Lomonosov Moscow State University, Moscow, Russia 2 Vladimir State University named after Alexander and Nikolay Stoletovs, Vladimir, Russia Correspondence: Trifonova Tatiana Anatolievna, Lomonosov Moscow State University, GSP-1, Leninskie Gory, Moscow, 119991, Russia. E-mail: [email protected] Received: October 15, 2014 Accepted: October 25, 2014 Online Published: December 7, 2014 doi:10.5539/mas.v9n2p78 URL: http://dx.doi.org/10.5539/mas.v9n2p78 Abstract The aim of this research is to analyze the demographic situation of the river Klyazma basin in the post-Soviet period and throughout the history of development and settlement of the region; identification of the major centres of vectors differentiation of population development in the examined basin; analysis of socio-economic, natural and environmental factors affecting the population distribution within the basin and demographic processes that are typical for the region. We must take into account that the population demographics of the region is largely dependent on diversity and state of resources of the area and their involvement into the economic sphere, however, it is obvious that the state of the economy is also a key factor in ensuring the sustainability of ecosystems and geo-systems of the river basin. The river Klyazma basin area has a long historical path of development and transformation of natural systems into natural and man-made landscape complexes.