WSSG Newsletter39
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Development of Forest Sector in the Arkhangelsk Oblast During the Transition Period of the 1990S
Development of forest sector in the Arkhangelsk oblast during the transition period of the 1990s ALBINA PASHKEVICH Pashkevich Albina (2003). Development of forest sector in the Arkhangelsk oblast during the transition period of the 1990s. Fennia 181: 1, pp. 13–24. Helsinki. ISSN 0015-0010. The Arkhangelsk oblast has long been one of Russia’s most important forest industrial regions. This paper analyses the changes in accessibility of forest resources and forest commodity production during the transition period in the 1990s. Special attention is given to firm restructuring, active roles of domestic capital and the different survival strategies that have been developed by in- dustries in the region. Further analysis deals with signs of economic recovery in the forest sector due to the processes of restructuring, modernisation and self-organisation. Albina Pashkevich, Spatial Modelling Centre (SMC), Department of Social and Economic Geography, Umeå University, Box 839, SE-98128 Kiruna, Sweden. E-mail: [email protected]. MS received 12 August 2002. Introduction adoption of a new. Some suggest that this proc- ess has been deeply embedded in the nature of The shift from central planning to a market-based the socialist system (Dingsdale 1999; Hamilton economy in Russia culminated with the dramatic 1999) and that the legacy of the communism has economic and political reorientation that began been only partly removed, and instead has mere- in the 1990s. This transition towards a market-ori- ly been reworked in a complex way (Smith 1997). ented and outward-looking economic system led Others say that reforms have actually ended the by private sector has created new challenges and old ‘command economy’ but have instead suc- opportunities. -

RCN #33 21/8/03 13:57 Page 1
RCN #33 21/8/03 13:57 Page 1 No. 33 Summer 2003 Special issue: The Transformation of Protected Areas in Russia A Ten-Year Review PROMOTING BIODIVERSITY CONSERVATION IN RUSSIA AND THROUGHOUT NORTHERN EURASIA RCN #33 21/8/03 13:57 Page 2 CONTENTS CONTENTS Voice from the Wild (Letter from the Editors)......................................1 Ten Years of Teaching and Learning in Bolshaya Kokshaga Zapovednik ...............................................................24 BY WAY OF AN INTRODUCTION The Formation of Regional Associations A Brief History of Modern Russian Nature Reserves..........................2 of Protected Areas........................................................................................................27 A Glossary of Russian Protected Areas...........................................................3 The Growth of Regional Nature Protection: A Case Study from the Orlovskaya Oblast ..............................................29 THE PAST TEN YEARS: Making Friends beyond Boundaries.............................................................30 TRENDS AND CASE STUDIES A Spotlight on Kerzhensky Zapovednik...................................................32 Geographic Development ........................................................................................5 Ecotourism in Protected Areas: Problems and Possibilities......34 Legal Developments in Nature Protection.................................................7 A LOOK TO THE FUTURE Financing Zapovedniks ...........................................................................................10 -

Transport and Infrastructural Basis of the Tourism Development Strategy in the Arkhangelsk Oblast © Aleksandr Yu
Aleksandr Yu. TSVETKOV. Transport and infrastructural basis … 35 UDC [338.48+332.14](470.11)(045) DOI: 10.37482/issn2221-2698.2020.38.44 Transport and infrastructural basis of the tourism development strategy in the Arkhangelsk Oblast © Aleksandr Yu. TSVETKOV, Cand. Sci. (Econ.), associate professor E-mail: [email protected] Department of Management, Higher School of Economics, Management and Law, Northern (Arctic) Federal University named after M.V. Lomonosov, Arkhangelsk, Russia Abstract. The article, devoted to the analysis of transport and geographical locations, describes possible strategies for the development of tourism in the Arkhangelsk Oblast. The main goal of the research was the development of logistic schemes of the transportation of tourists from the places of formation of tourist flows to the Arkhangelsk Oblast. The methodological basis of the article is to determine the economic dis- tances between potential tourist distribution centers and their places of interest in the area. Moscow, St. Petersburg, and Arkhangelsk were considered as the main towns of departure. Kargopol, Solvychegodsk, Kholmogory and Lomonosovo, Solovki, Kenozersky National Park, and Pinega caves are regarded as the main sites of tourist interest in the Arkhangelsk Oblast. It was determined that Kargopol is the most acces- sible for tourists, and Kenozersky National Park is the most recognizable by tourists but the least accessible. The object of world cultural heritage, the Monastery of the Transfiguration of the Saviour on Solovki is the most accessible for tourists from the territory of Karelia. It is recommended to optimize the schedule and to synchronize the work of transport for tourists to improve the transport accessibility of recreational facili- ties in the area. -

Download Article (PDF)
Advances in Social Science, Education and Humanities Research, volume 144 3rd International Conference on Arts, Design and Contemporary Education (ICADCE 2017) Rituals and Customs as a Reflection of the Folk Tradition The Russian North in the Context of Culture Magda Djichonaya Institute of Slavic Culture Russian State University named after A.N. Kosygin Moscow, Russia E-mail: [email protected] Abstract—The article is devoted to folklore traditions and is II. TRADITIONAL HOUSEHOLD WAY OF LIFE OF considered by the example of the Russian North. Ethno- INHABITANTS OF THE ARKHANGELSK REGION cultural realities reflect the specifics of the structure of artistic culture and of the existence in it of authentic folklore. Russia‘s Arkhangelsk Region, for instance, due to its remoteness, is distinguished by preserved in the area unique Keywords—polyfunctionality of art; art in the context of centuries-old folk traditions. Its art presents the masterpiece culture; folk traditions; ethnic and cultural realities of people and the perception in the mass. Folklore traditions are very well preserved, as handed down from generation to I. INTRODUCTION generation. The specificity of the cultural and logical approach to the Description of nature, navigation, hunting, fishing, study of art lies in its consideration as part of a holistic socio- peculiarities of female labor, buildings, clothing, etc. cultural space as a special sphere of human activity and, contained numerous realities of everyday life of the fishing finally, as a cultural phenomenon in which the artistic and population of the Russian North, from which came the aesthetic values are accumulated. storytellers. Openness to artistic culture as a system lies in its ability The key elements of the way of life of northerners — to restrict and expand one‘s frame of graduation. -
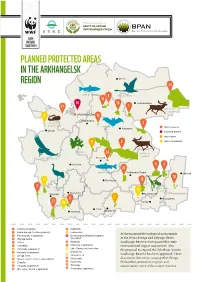
Planned Protected Areas in the Arkhangelsk Region
PLANNED PROTECTED AREAS IN THE ARKHANGELSK Mezen REGION 23 atlantic salmon 8 whooper swan 13 16 14 Leshukonskoe 17 1 wild forest reindeer ARKHANGELSK 9 Новодвинск 12 Kholmogory freshwater 3 pearl mussel вrandt's nature reserve bat Karpogory Onega botanical garden nature park 11 2 nature monument tree lungwort 24 wild forest reindeer white-tailed eagle Bereznik Plesetsk 4 4 22 atlantic salmon Verkhnyaya Toyma Yarensk 10 15 Shenkursk 18 5 common lady’s slipper crane 19 orchid Nyandoma Kargopol Krasnoborsk 7 6 Koryazhma 21 20 Котлас Velsk Konosha Oktyabrsky 1 Verkhnechelassky 14 Sebboloto 2 15 Dvina-Pinega (Verkhneyulovsky) Lekshmokh At the moment the ecological assessments 3 Puchkomsky (expansion) 16 Dendrological (botanical) garden 4 Uftyuga-Ilesha “SevNIILH” of the Dvina-Pinega and Uftyuga-Ilesha 5 Atleka 17 Solzinsky Landscape Reserves have passed the state 6 Voloshsky 18 Shilovsky (expansion) environmental impact assessment. Also 7 Ustiansky (expansion) 19 Lake Churozero (correction the proposal to expand the Zelezhnye Vorota 8 Kuloysky (expansion) of borders) 20 Landscape Reserve has been approved. These 9 Onega coast Turovsky Les 21 10 Nature reserve in the Lensky district Kovzhinsky documents determine, among other things, 22 11 Zvozsky Soyginsky the borders, protection regimes and 23 12 Chugsky (expansion) Timansky conservation zones of the nature reserves. 24 13 Zheleznye Vorota (expansion) Klonovsky (expansion) Intact forest landscapes are Arkhangelsk Region has been shaped by optimal habitats for such coniferous forests, covering most of its animals as bears, pine martens, elks and territory. Its virgin forests, which have not capercaillies. been disturbed by human activities, play a key role in protecting populations of rare and threatened species. -

Building the Landysh in the Russian Far East
CRISTINA CHUEN & TAMARA TROYAKOVA Report The Complex Politics of Foreign Assistance: Building the Landysh in the Russian Far East CRISTINA CHUEN & TAMARA TROYAKOVA Cristina Chuen is a Research Associate at the Center for Nonproliferation Studies(CNS), Monterey Institute of International Studies, and a Ph.D. candidate in International Affairs at the University of California at San Diego, specializing in local government and center-region relations in Russia and China. Her recent publications include the co-authored “Russia’s Blue Water Blues,” in the Bulletin of the Atomic Scientists (January 2001), and an editorial in the International Herald Tribune on nuclear regulation in Russia. Tamara Troyakova is a Senior Researcher at the Institute of History, Archaeology, and Ethnography of Peoples of the Far East, Russian Academy of Sciences, in Vladivostok, Russia. She also served as Associate Professor at the Far Eastern State University (1995-1999) and the Far Eastern Commercial Institute (1992-1994) in Vladivostok. In addition, Ms. Troyakova was Visiting Fellow at CNS in 2000 and a Visiting Scholar at the George F. Kennan Institute for Advanced Russian Studies, Woodrow Wilson International Center for Scholars, in 1999. his report, based on extensive interviews and con- dismantle though the Cooperative Threat Reduction Pro- temporaneous news accounts, examines the poli- gram. Of the remaining 143 submarines, 87 still have tics surrounding the construction of a Japanese- nuclear fuel aboard, while the other 56 have been defueled T 1 financed liquid radioactive waste (LRW) treatment plant but not dismantled. These vessels pose a global prolif- in the Russian Far East. The facility removes radioactive eration threat, due to the large amounts of highly enriched contamination from water that has circulated in subma- uranium (HEU)—a key ingredient of nuclear weapons— rine nuclear reactors as coolant. -

Meeting Places and Social Capital Supporting Rural Landscape Stewardship: a Pan-European Horizon Scanning
Copyright © 2021 by the author(s). Published here under license by the Resilience Alliance. Angelstam, P., M. Fedoriak, F. Cruz, J. Muñoz-Rojas, T. Yamelynets, M. Manton, C.-L. Washbourne, D. Dobrynin, Z. Izakovičova, N. Jansson, B. Jaroszewicz, R. Kanka, M. Kavtarishvili, L. Kopperoinen, M. Lazdinis, M. J. Metzger, D. Özüt, D. Pavloska Gjorgjieska, F. J. Sijtsma, N. Stryamets, A. Tolunay, T. Turkoglu, B. Van der Moolen, A. Zagidullina, and A. Zhuk. 2021. Meeting places and social capital supporting rural landscape stewardship: A Pan-European horizon scanning. Ecology and Society 26(1):11. https://doi.org/10.5751/ES-12110-260111 Research Meeting places and social capital supporting rural landscape stewardship: A Pan-European horizon scanning Per Angelstam 1, Mariia Fedoriak 2, Fatima Cruz 3, José Muñoz-Rojas 4, Taras Yamelynets 5, Michael Manton 6, Carla-Leanne Washbourne 7, Denis Dobrynin 8, Zita Izakovičova 9, Nicklas Jansson 10, Bogdan Jaroszewicz 11, Robert Kanka 9, Marika Kavtarishvili 12, Leena Kopperoinen 13, Marius Lazdinis 14, Marc J. Metzger 15, Deniz Özüt 16, Dori Pavloska Gjorgjieska 17, Frans J. Sijtsma 18, Nataliya Stryamets 19,20, Ahmet Tolunay 21, Turkay Turkoglu 22, Bert van der Moolen 23, Asiya Zagidullina 24 and Alina Zhuk 25 ABSTRACT. Achieving sustainable development as an inclusive societal process in rural landscapes, and sustainability in terms of functional green infrastructures for biodiversity conservation and ecosystem services, are wicked challenges. Competing claims from various sectors call for evidence-based adaptive collaborative governance. Leveraging such approaches requires maintenance of several forms of social interactions and capitals. Focusing on Pan-European regions with different environmental histories and cultures, we estimate the state and trends of two groups of factors underpinning rural landscape stewardship, namely, (1) traditional rural landscape and novel face-to-face as well as virtual fora for social interaction, and (2) bonding, bridging, and linking forms of social capital. -

Supplement of Geosci
Supplement of Geosci. Model Dev., 11, 1343–1375, 2018 https://doi.org/10.5194/gmd-11-1343-2018-supplement © Author(s) 2018. This work is distributed under the Creative Commons Attribution 4.0 License. Supplement of LPJmL4 – a dynamic global vegetation model with managed land – Part 1: Model description Sibyll Schaphoff et al. Correspondence to: Sibyll Schaphoff ([email protected]) The copyright of individual parts of the supplement might differ from the CC BY 4.0 License. S1 Supplementary informations to the evaluation of the LPJmL4 model The here provided supplementary informations give more details to the evaluations given in Schaphoff et al.(under Revision). All sources and data used are described in detail there. Here we present ad- ditional figures for evaluating the LPJmL4 model on a plot scale for water and carbon fluxes Fig. S1 5 - S16. Here we use the standard input as described by Schaphoff et al.(under Revision, Section 2.1). Furthermore, we evaluate the model performance on eddy flux tower sites by using site specific me- teorological input data provided by http://fluxnet.fluxdata.org/data/la-thuile-dataset/(ORNL DAAC, 2011). Here the long time spin up of 5000 years was made with the input data described in Schaphoff et al.(under Revision), but an additional spin up of 30 years was conducted with the site specific 10 input data followed by the transient run given by the observation period. Comparisons are shown for some illustrative stations for net ecosystem exchange (NEE) in Fig. S17 and for evapotranspira- tion Fig. S18. -

'Sustainable Forestry' in Russia Is Destroying the Great Northern Forest
EYE ON THE TAIGA HOW INDUSTRY’S CLAIMED ‘SUSTAINABLE FORESTRY’ IN RUSSIA IS DESTROYING THE GREAT NORTHERN FOREST EYE ON THE TIAGA 1 The timber sector in Arkhangelsk Oblast is still heavily reliant on the logging of Intact Forest Landscapes to maintain the large inputs of softwood timber needed to feed the increasing demand from the area’s expanding sawmills and pulp mills. Cover © Greenpeace Contents page © Igor Podgorny / Greenpeace 2 EYE ON THE TIAGA CONTENTS EYE ON THE TAIGA: HOW INDUSTRY’S CLAIMED ‘SUSTAINABLE FORESTRY’ IN RUSSIA IS DESTROYING THE GREAT NORTHERN FOREST 1 SUMMARY 4 SECTION 1: GLOBAL CONTEXT TO PROTECTING THE GREAT NORTHERN FOREST 8 SECTION 2: INDUSTRY’S CONTINUED DEPENDENCE ON INTACT FOREST LANDSCAPE DESTRUCTION 16 SECTION 3: THE BATTLE TO PROTECT THE DVINSKY FOREST – AN IFL BIODIVERSITY HOTSPOT 20 SECTION 4: MARKETS FOR KEY INDUSTRY PLAYERS IN ARKHANGELSK OBLAST 26 SECTION 5: WILL FOREST CERTIFICATION PROTECT IFLS IN ARKHANGELSK? 28 REPORT CONCLUSIONS 30 GREENPEACE DEMANDS 31 ACRONYMS AND BIBLIOGRAPHY 34 ENDNOTES 37 EYE ON THE TIAGA 3 SUMMARY The scale of this crisis in the Great Northern Forest can be judged from fact that this rate of IFL loss was over one-and- a-half times the average annual rate of deforestation in the Amazon rainforest for most of the same period. THE EXTINCTION contribute towards protecting at least change than other forests.8 It is home to CRISIS – 17% of the world’s terrestrial areas a rich diversity of native mammals ranging INTERNATIONAL important for biodiversity and ecosystem from elk and deer, through beavers, services (including primary forests) by wolverines and porcupines, to martens, COMMITMENTS TO means of ‘ecologically representative and squirrels and lemmings. -

73 Escoube,2015.Pdf
C. R. Geoscience 347 (2015) 377–385 Contents lists available at ScienceDirect Comptes Rendus Geoscience ww w.sciencedirect.com Hydrology, Environment (Surface Geochemistry) Iron isotope systematics in Arctic rivers a,b,i,1 b,c, d,e,f Raphaelle Escoube , Olivier J. Rouxel *, Oleg S. Pokrovsky , g h a Andrew Schroth , Robert Max Holmes , Olivier F.X. Donard a LCABIE, Universite´ de Pau et des pays de l’Adour, CNRS UMR 525, He´lioparc, 64053 Pau, France b Department of Marine Chemistry and Geochemistry, Woods Hole Oceanographic Institution, MA 02543 Woods Hole, USA c IFREMER, REM/GM, Centre de Brest, Plouzane´, France d Georesources and Environment Toulouse GET, UMR 5563, CNRS, Universite´ Paul-Sabatier, 31400 Toulouse, France e Institute of Ecological Problems of the North, Ural Branch RAS, Arkhangelsk, Russia f BIO-GEO-CLIM Laboratory, Tomsk State University, Tomsk, Russia g Department of Geology, University of Vermont, VT05405 Burlington, USA h Woods Hole Research Center, 02540 Falmouth, MA, USA i Universita¨t zu Ko¨ln, Institut fu¨r Geologie und Mineralogie, Zu¨lpicher Str. 49b, 50674 Ko¨ln, Germany A R T I C L E I N F O A B S T R A C T Article history: The input of iron to the Arctic Ocean plays a critical role in the productivity of aquatic Received 8 February 2015 ecosystems and is potentially impacted by climate change. We examine Fe isotope Accepted after revision 23 April 2015 systematics of dissolved and colloidal Fe from several Arctic and sub-Arctic rivers in Available online 2 July 2015 56 northern Eurasia and Alaska. -

Bird Communities in European Taiga Forest: a Comparison Between A
Bird communities in European taiga forest: A comparison between a large forest block in Archangelsk, Russia, and some small-grained old-growth fragments in central Norway Per Gustav Thingstad1, Ole Jakob Sørensen2 & Vladimir Naumov3 Thingstad, P.G., Sørensen, O.J. & Naumov, V. 2006. Bird communities in European taiga forest: A comparison between a large forest block in Archangelsk, Russia, and some small-grained old-growth fragments in central Norway. - Ornis Norvegica 29: 46-58. The large intact areas of old-growth forest of the Archangelsk oblast represent today a possibility of studying taiga ecology in ecosystems that we do not find in Fennoscandia. They are to be regarded as the sources for the taiga-elements in our own coniferous forests. This study of bird communities was done in the core of one of the old remaining, intact forests of Archangelsk, alongside the Yula river in Pinega and Vinogradovsky rayons («regions»). Bird communities in mature spruce and pine dominated forests were studied in spring 2005. These communities from the core area of the taiga are compared with coniferous forest in the rural municipality of Lierne in eastern central Norway, studied in 2004. This latter area should be representative for the westernmost parts of the taiga. Due to extensive forestry exploitation and natural heterogeneity it is heavily fragmented, leav- ing behind only smaller fragments of more or less coherent old-growth stands. Findings in our comparison: • Species dependent on sufficient amount of dead wood, e.g. woodpeckers, comprise far less of the bird com- munities within the small-grained old-growth fragments in Lierne compared with the situation in the virgin taiga at Yula. -

North Russia: 1918-1919
MASTER'S THESIS M-2208 BALDWIN, Robert Cushman THE ALLIED MILITARY EXPEDITION TO NORTH RUSSIA: 1918-1919. The American University, M.A., 1969 History, modern University Microfilms, A XEROX Company , Ann Arbor, Michigan THIS DISSERTATION HAS BEEN MICROFILMED EXACTLY AS RECEIVED Reproduced with permission of the copyright owner. Further reproduction prohibited without permission. THE ALLIED MILITARY EXPEDITION TO NORTH RUSSIA, 1918-1919 by Robert Cushman Baldwin Submitted to the Faculty of the College of Arts and Sciences of the American University in Partial Fulfillment of the Requirements for the Degree of Master of Arts in History Signatures of Committee: Chairman, Dean of the Go^ege Date, /. /fJI___ Date,. June 1969 THE AMERICAN UNIVERSITY The American University JAN 27 1970 Washington, D. C. W7 Reproduced with permission of the copyright owner. Further reproduction prohibited without permission. ABSTRACT The Allied military expedition to North Russia in 1918 is an historical event unknown to most Americans. However, Soviet historiography on the subject is extensive and at least one Soviet leader has cited the event as a fundamental cause of the consistently poor relations between the Soviet Union and the Western Powers. This paper seeks to examine the reasons for the expedition and to evaluate its effect. The records of the Supreme War Council of the Allied Powers were examined to determine the military reasons advanced for dispatching the expedition, and the records of the American contingent were reviewed to develop the record of the operations in which it engaged. The conclusion of this thesis is that the Allied anabasis failed to attain any of the diverse objectives set for it and that, because military force was applied without a clear-cut central purpose, the expedition was an utter failure.