A Study of the Renal Circulation, with Spec Ia L
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Urinary System
URINARY SYSTEM Ján Líška DVM, PhD Institut of Histology and Embryology, Faculty of Medicine, Comenius University Urinary system • The kidneys are the organ with multiple functions: • filtration of the blood • excretion of metabolic waste products and related removal of toxins • maintenance blood volume • regulation of acid-base balance • regulation of fluid and electrolyte balance • production of the hormones The other components of urinary system are accessory. Their function is essentially in order to eliminate urine. Urinary system - anatomy • Kidney are located in the retroperitoneal space • The surface of the kidney is covered by a fibrous capsule of dense connective tissue. • This capsule is coated with adipose capsule. • Each kidney is attached to a ureter, which carries urine to the bladder and urine is discharged out through the urethra. ANATOMIC STRUCTURE OF THE KIDNEY RENAL LOBES CORTEX outer shell columns Excretory portion medullary rays MEDULLA medullary pyramids HILUM Collecting system blood vessels lymph vessels major calyces nerves RENAL PELVIS minor calyces ureter Cortex is the outer layer surrounding the internal medulla. The cortex contains renal corpuscles, convoluted parts of prox. and dist. tubules. Renal column: the renal tissue projection between two medullary pyramids which supports the cortex. Renal pyramids: the conical segments within the medulla. They contain the ductal apparatus and stright parts of the tubules. They posses papilla - having openings through which urine passes into the calyces. Each pyramid together with the associated overlying cortex forms a renal lobe. renal pyramid papilla minor calix minor calyx Medullary rays: are in the middle of cortical part of the renal lobe, consisting of a group of the straight portiones of nephrons and the collec- medullary rays ting tubules (only straight tubules). -

The Renal Vascular System of the Monkey: a Gross Anatomical Description MARK J
J. Anat. (1987), 153, pp. 123-137 123 With 8 figures Printed in Great Britain The renal vascular system of the monkey: a gross anatomical description MARK J. HORACEK, ALVIN M. EARLE AND JOSEPH P. GILMORE Departments ofAnatomy and Physiology and Biophysics, University of Nebraska College of Medicine, Omaha, Nebraska 68105, U.S.A. (Accepted 12 September 1986) INTRODUCTION Monkeys are frequently used as a model for physiological experiments involving the kidney. It is known that physiological disparity between mammalian species can often be elucidated by a study of the structural differences between these species. Despite this fact, the monkey's renal vasculature has not been described in sufficient detail. Such a description would be useful since monkey experimentation plays an important role in understanding human physiology. Also, the branching pattern and segmen- tation of the monkey's renal vasculature is interesting from both comparative and experimental viewpoints. Fourman & Moffat (1971) indicated that although all mammalian kidneys are somewhat similar, there are several species-specific differences in terms oforganisation, microstructure and function. They conducted several extensive vascular studies on mammals, including several rodents and carnivores, but did not include any specific information concerning the monkey. Graves (1971) and Fine & Keen (1966) have described the branching patterns and segmentation of human kidneys. However, comparable information pertaining to the monkey kidney is not available. The purpose of the present study is to provide a detailed morphological description of the gross renal vasculature in the kidneys of two species of monkeys, Macacafascicularis and Macaca mulatta. MATERIALS AND METHODS Twelve monkeys (Macacafascicularis and Macaca mulatta) were used in this study after they had been utilised for electrophysiological experimentation. -

Anatomy and Physiology of the Bowel and Urinary Systems
PMS1 1/26/05 10:52 AM Page 1 Anatomy and Physiology of the Bowel and 1 Urinary Systems Anthony McGrath INTRODUCTION The aim of this chapter is to increase the reader’s under- standing of the small and large bowel and urinary system as this will enhance their knowledge base and allow them to apply this knowledge when caring for patients who are to undergo stoma formation. LEARNING OBJECTIVES By the end of this chapter the reader will have: ❏ an understanding of the anatomy and physiology of the small and large bowel; ❏ an understanding of the anatomy and physiology of the urinary system. GASTROINTESTINAL TRACT The gastrointestinal (GI) tract (Fig. 1.1) consists of the mouth, pharynx, oesophagus, stomach, duodenum, jejunum, small and large intestines, rectum and anal canal. It is a muscular tube, approximately 9m in length, and it is controlled by the autonomic nervous system. However, while giving a brief outline of the whole system and its makeup, this chapter will focus on the anatomy and physiology of the small and large bowel and the urinary system. The GI tract is responsible for the breakdown, digestion and absorption of food, and the removal of solid waste in the form of faeces from the body. As food is eaten, it passes through each section of the GI tract and is subjected to the action of various 1 PMS1 1/26/05 10:52 AM Page 2 1 Anatomy and Physiology of the Bowel and Urinary Systems Fig. 1.1 The digestive system. Reproduced with kind permission of Coloplast Ltd from An Introduction to Stoma Care 2000 2 PMS1 1/26/05 10:52 AM Page 3 Gastrointestinal Tract 1 digestive fluids and enzymes (Lehne 1998). -
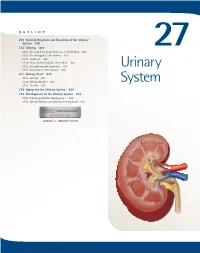
Urinary System
OUTLINE 27.1 General Structure and Functions of the Urinary System 818 27.2 Kidneys 820 27 27.2a Gross and Sectional Anatomy of the Kidney 820 27.2b Blood Supply to the Kidney 821 27.2c Nephrons 824 27.2d How Tubular Fluid Becomes Urine 828 27.2e Juxtaglomerular Apparatus 828 Urinary 27.2f Innervation of the Kidney 828 27.3 Urinary Tract 829 27.3a Ureters 829 27.3b Urinary Bladder 830 System 27.3c Urethra 833 27.4 Aging and the Urinary System 834 27.5 Development of the Urinary System 835 27.5a Kidney and Ureter Development 835 27.5b Urinary Bladder and Urethra Development 835 MODULE 13: URINARY SYSTEM mck78097_ch27_817-841.indd 817 2/25/11 2:24 PM 818 Chapter Twenty-Seven Urinary System n the course of carrying out their specific functions, the cells Besides removing waste products from the bloodstream, the uri- I of all body systems produce waste products, and these waste nary system performs many other functions, including the following: products end up in the bloodstream. In this case, the bloodstream is ■ Storage of urine. Urine is produced continuously, but analogous to a river that supplies drinking water to a nearby town. it would be quite inconvenient if we were constantly The river water may become polluted with sediment, animal waste, excreting urine. The urinary bladder is an expandable, and motorboat fuel—but the town has a water treatment plant that muscular sac that can store as much as 1 liter of urine. removes these waste products and makes the water safe to drink. -

The Urinary System Dr
The urinary System Dr. Ali Ebneshahidi Functions of the Urinary System • Excretion – removal of waste material from the blood plasma and the disposal of this waste in the urine. • Elimination – removal of waste from other organ systems - from digestive system – undigested food, water, salt, ions, and drugs. + - from respiratory system – CO2,H , water, toxins. - from skin – water, NaCl, nitrogenous wastes (urea , uric acid, ammonia, creatinine). • Water balance -- kidney tubules regulate water reabsorption and urine concentration. • regulation of PH, volume, and composition of body fluids. • production of Erythropoietin for hematopoieseis, and renin for blood pressure regulation. Anatomy of the Urinary System Gross anatomy: • kidneys – a pair of bean – shaped organs located retroperitoneally, responsible for blood filtering and urine formation. • Renal capsule – a layer of fibrous connective tissue covering the kidneys. • Renal cortex – outer region of the kidneys where most nephrons is located. • Renal medulla – inner region of the kidneys where some nephrons is located, also where urine is collected to be excreted outward. • Renal calyx – duct – like sections of renal medulla for collecting urine from nephrons and direct urine into renal pelvis. • Renal pyramid – connective tissues in the renal medulla binding various structures together. • Renal pelvis – central urine collecting area of renal medulla. • Hilum (or hilus) – concave notch of kidneys where renal artery, renal vein, urethra, nerves, and lymphatic vessels converge. • Ureter – a tubule that transport urine (mainly by peristalsis) from the kidney to the urinary bladder. • Urinary bladder – a spherical storage organ that contains up to 400 ml of urine. • Urethra – a tubule that excretes urine out of the urinary bladder to the outside, through the urethral orifice. -

Normal Vascular and Glomerular Anatomy
Normal Vascular and Glomerular Anatomy Arthur H. Cohen Richard J. Glassock he topic of normal vascular and glomerular anatomy is intro- duced here to serve as a reference point for later illustrations of Tdisease-specific alterations in morphology. CHAPTER 1 1.2 Glomerulonephritis and Vasculitis FIGURE 1-1 A, The major renal circulation. The renal artery divides into the interlobar arteries (usually 4 or 5 divisions) that then branch into arcuate arteries encompassing the corticomedullary Interlobar junction of each renal pyramid. The interlobular arteries (multiple) originate from the artery arcuate arteries. B, The renal microcirculation. The afferent arterioles branch from the interlobular arteries and form the glomerular capillaries (hemi-arterioles). Efferent arteri- Arcuate oles then reform and collect to form the post-glomerular circulation (peritubular capillar- artery Renal ies, venules and renal veins [not shown]). The efferent arterioles at the corticomedullary artery junction dip deep into the medulla to form the vasa recta, which embrace the collecting tubules and form hairpin loops. (Courtesy of Arthur Cohen, MD.) Pyramid Pelvis Interlobular Ureter artery A Afferent arteriole Interlobular artery Glomerulus Arcuate artery Efferent arteriole Collecting tubule Interlobar artery B Normal Vascular and Glomerular Anatomy 1.3 FIGURE 1-2 (see Color Plate) Microscopic view of the normal vascular and glomerular anatomy. The largest intrarenal arteries (interlobar) enter the kidneys between adjacent lobes and extend toward the cortex on the side of a pyramid. These arteries branch dichotomously at the corti- comedullary junction, forming arcuate arteries that course between the cortex and medulla. The arcuate arteries branch into a series of aa ILA interlobular arteries that course at roughly right angles through the cortex toward the capsule. -

Kidney Structure Renal Lobe Renal Lobule
Kidney Structure Capsule Hilum • ureter → renal pelvis → major and minor calyxes • renal artery and vein → segmental arteries → interlobar arteries → arcuate arteries → interlobular arteries Medulla • renal pyramids • cortical/renal columns Cortex • renal corpuscles • cortical labryinth of tubules • medullary rays Renal Lobe Renal Lobule = renal pyramid & overlying cortex = medullary ray & surrounding cortical labryinth Cortex Medulla Papilla Calyx Sobotta & Hammersen: Histology 1 Uriniferous Tubule Nephron + Collecting tubule Nephron Renal corpuscle produces glomerular ultrafiltrate from blood Ultrafiltrate is concentrated • Proximal tubule • convoluted • straight • Henle’s loop • thick descending • thin • thick ascending • Distal tubule • Collecting tubule Juxtaglomerular apparatus • macula densa in distal tubule •JG cells in afferent arteriole •extraglomerular mesangial cells Glomerulus • fenestrated capillaries • podocytes • intraglomerular mesangial cells 2 Urinary Filtration Urinary Membrane Membrane Podocytes • Endothelial cell • 70-90 nm fenestra restrict proteins > 70kd • Basal lamina • heparan sulfate is negatively charged • produced by endothelial cells & podocytes • phagocytosed by mesangial cells • Podocytes • pedicels 20-40 nm apart • diaphragm 6 nm thick with 3-5 nm slits • podocalyxin in glycocalyx is negatively charged 3 Juxtaglomerular Apparatus Macula densa in distal tubule • monitor Na+ content and volume in DT • low Na+: • stimulates JG cells to secrete renin • stimulates JG cells to dilate afferent arteriole • tall, -

(A) Adrenal Gland Inferior Vena Cava Iliac Crest Ureter Urinary Bladder
Hepatic veins (cut) Inferior vena cava Adrenal gland Renal artery Renal hilum Aorta Renal vein Kidney Iliac crest Ureter Rectum (cut) Uterus (part of female Urinary reproductive bladder system) Urethra (a) © 2018 Pearson Education, Inc. 1 12th rib (b) © 2018 Pearson Education, Inc. 2 Renal cortex Renal column Major calyx Minor calyx Renal pyramid (a) © 2018 Pearson Education, Inc. 3 Cortical radiate vein Cortical radiate artery Renal cortex Arcuate vein Arcuate artery Renal column Interlobar vein Interlobar artery Segmental arteries Renal vein Renal artery Minor calyx Renal pelvis Major calyx Renal Ureter pyramid Fibrous capsule (b) © 2018 Pearson Education, Inc. 4 Cortical nephron Fibrous capsule Renal cortex Collecting duct Renal medulla Renal Proximal Renal pelvis cortex convoluted tubule Glomerulus Juxtamedullary Ureter Distal convoluted tubule nephron Nephron loop Renal medulla (a) © 2018 Pearson Education, Inc. 5 Proximal convoluted Peritubular tubule (PCT) Glomerular capillaries capillaries Distal convoluted tubule Glomerular (DCT) (Bowman’s) capsule Efferent arteriole Afferent arteriole Cells of the juxtaglomerular apparatus Cortical radiate artery Arcuate artery Arcuate vein Cortical radiate vein Collecting duct Nephron loop (b) © 2018 Pearson Education, Inc. 6 Glomerular PCT capsular space Glomerular capillary covered by podocytes Efferent arteriole Afferent arteriole (c) © 2018 Pearson Education, Inc. 7 Filtration slits Podocyte cell body Foot processes (d) © 2018 Pearson Education, Inc. 8 Afferent arteriole Glomerular capillaries Efferent Cortical arteriole radiate artery Glomerular 1 capsule Three major renal processes: Rest of renal tubule 11 Glomerular filtration: Water and solutes containing smaller than proteins are forced through the filtrate capillary walls and pores of the glomerular capsule into the renal tubule. Peritubular 2 capillary 2 Tubular reabsorption: Water, glucose, amino acids, and needed ions are 3 transported out of the filtrate into the tubule cells and then enter the capillary blood. -

The Distal Convoluted Tubule and Collecting Duct
Chapter 23 *Lecture PowerPoint The Urinary System *See separate FlexArt PowerPoint slides for all figures and tables preinserted into PowerPoint without notes. Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Introduction • Urinary system rids the body of waste products. • The urinary system is closely associated with the reproductive system – Shared embryonic development and adult anatomical relationship – Collectively called the urogenital (UG) system 23-2 Functions of the Urinary System • Expected Learning Outcomes – Name and locate the organs of the urinary system. – List several functions of the kidneys in addition to urine formation. – Name the major nitrogenous wastes and identify their sources. – Define excretion and identify the systems that excrete wastes. 23-3 Functions of the Urinary System Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Diaphragm 11th and 12th ribs Adrenal gland Renal artery Renal vein Kidney Vertebra L2 Aorta Inferior vena cava Ureter Urinary bladder Urethra Figure 23.1a,b (a) Anterior view (b) Posterior view • Urinary system consists of six organs: two kidneys, two ureters, urinary bladder, and urethra 23-4 Functions of the Kidneys • Filters blood plasma, separates waste from useful chemicals, returns useful substances to blood, eliminates wastes • Regulate blood volume and pressure by eliminating or conserving water • Regulate the osmolarity of the body fluids by controlling the relative amounts of water and solutes -

Renal Doppler Complete (Renal Artery Stenosis)
UT Southwestern Department of Radiology Ultrasound – Renal Doppler Complete (Renal Artery Stenosis) PURPOSE: To evaluate the vasculature associated with the native kidneys for arterial stenosis. SCOPE: Applies to all ultrasound abdominal studies performed in Imaging Services / Radiology EPIC ORDERABLE: • US Doppler Renal (perform this protocol only) CHARGEABLES: • US Doppler Renal (US DOPPLER COMPLETE CPT 93975) • Add to US Renal if ordered together (US RENAL COMPLETE, CPT 76770) o See specific US Renal protocol for details regarding a complete renal examination. INDICATIONS: • Suspected renal artery stenosis (examples: hypertension resistive to treatment, hypertension in young patients etc.); • Suspected vasculitis or fibromuscular dysplasia (FMD); • Abnormal findings on other imaging studies; • If renal vessel patency is the clinical concern, refer to protocol “US Renal Doppler Patency”; • If nutcracker Syndrome (renal vein entrapment) is the clinical concern, refer to protocol “US Renal Doppler Nutcracker). CONTRAINDICATIONS: • No absolute contraindications • For unstable, hospitalized, and ventilated patients, ordering providers should consider alternative modalities for ruling out RAS. Doppler is frequently nondiagnostic in these situations. A diagnostic Doppler exam for RAS should be reserved for stable outpatients who are able to comply with breathing instructions. EQUIPMENT: • Curvilinear transducer with a frequency range of 1-9 MHz that allows for appropriate penetration and resolution depending on patient’s body habitus PATIENT -

Anatomy of the Kidney
Anatomy of the kidney Renal block-Anatomy-Lecture 1 Editing file Color guide : Only in boys slides in Green Objectives Only in girls slides in Purple important in Red Notes in Grey By the end of this course you should be able to discuss : ● Components of the urinary system ● Kidney : 1. Shape & Position 2. Surface anatomy 3. External features 4. Hilum & its contents 5. Relation 6. Internal features 7. Blood supply 8. Lymph drainage 9. Nerve supply Introduction 3 ● Every day, each kidney filters liters (around 150 L per day ) of fluid from the bloodstream. ● Although the lungs and the skin also play roles in excretion, The kidneys bear the major responsibility for eliminating nitrogenous (nitrogen-containing) wastes, toxins, and drugs from the body. Excretes most of the Maintain acid-base waste products of balance of the blood. metabolism. By Erythropoietin Controls hormone stimulates Function of water & electrolyte bone marrow for RBCs kidney balance of the body. formation. Converts By Rennin vitamin D to its enzyme regulates active form. the blood pressure. The kidney : ● Kidneys are reddish brown in color. 4 ● Lie behind the peritoneum (retroperitoneal) on the posterior abdominal wall on either side of the vertebral column. ● They are largely under cover of the costal margin. kidney lies between T12-L3 ● With contraction of the diaphragm (during inspiration) the kidney moves downward as much as 2.5 cm. T12 Comparison between : L3 Right kidney Left kidney Anterior view lies slightly lower than the left due Location to the large size of the right lobe of Upper than the right the liver. -
Urinary System
Urinary system Sándor Katz M.D.,Ph.D. Urinary system - constituents • kidneys • ureters • urinary bladder • urethra Kidney Weight: 130-140g Kidneys - location 1. On the posterior body wall 2. Posterior to parietal peritoneum – retroperitoneal organ 3. At the level of T12-L2 (left kidney) and L1-L3 (right kidney) Kidneys - location Kidneys – covering structures 1. Renal (Gerota’s) fascia 2. Adipose capsule 3. Fibrous capsule Kidneys - neighbouring organs and structures Kidney – gross anatomy External structures: Hilum of kidney: 1. Renal vein 2. Renal artery 3. Ureter Internal structures: 1. Cortex 2. Medulla 3. Minor calyces 4. Major calyces 5. Renal pelvis Renal cortex Renal columns (Bertini’s columns) Renal medulla – renal pyramids A p p r o x i m a t e l y 3 0 pyramids are in each kidney and many of them are fused together. renal papilla Minor calyces 8-9 in each kidney Major calyces Approx. 3 in each kidney Renal pelvis Renal hilum - L1/L2 level renal sinus From anterior to posterior direction: 1. renal vein 2. renal artery 3. ureter From superior to inferior direction: 1. renal artery 2. renal vein 3. ureter Renal arteries - L1 level Renal artery • segmental arteries • interlobar arteries • arcuate arteries • interlobular arteries • afferent arterioles Renal veins left renal vein is longer than the right one and crosses over the aorta Renal veins right renal vein left renal vein is longer than the right one and crosses over the aorta left renal vein Tributaries of the renal veins • (stellate veins – only under the fibrous capsule) • interlobular veins • arcuate veins • interlobar veins • segmental veins Renal veins left suprarenal vein (empties into the left renal vein) left gonadal (testicular or ovarian) vein (empties into the left renal vein) The right suprarenal and gonadal veins empty into the IVC.