Phd Write-Up
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ken Matthews: 1934-2019
KEN MATTHEWS: 1934-2019 The world’s racewalking community was saddened in June 2019 to hear of the passing of Ken Matthews, Great Britain’s last surviving Olympic race walking Gold medallist. His death occurred on the evening of Sunday 2 nd June in Wrexham where he was a hospital in-patient. Kenneth ("Ken") Joseph Matthews was born on 21 June 1934 in Birmingham and started his race walk career as an 18- year-old, following in the footsteps of his father, Joe, who was a founding member of the now defunct Royal Sutton Coldfield Walking Club. Throughout his athletics career, Ken remained Midlands based, and remained a loyal member of Royal Sutton Coldfield Walking Club. An electrical maintenance engineer at a power station near his hometown of Sutton Coldfield, he became one of England's most successful ever racewalkers and dominated the world stage throughout the early 1960's. He won 17 national titles, as well as Olympic and European gold and, between 1964 to 1971 he held every British record from 5 miles to 2 hours, including a 10-mile world best of 69:40.6. Perhaps surprisingly, he did not dominate as a youngster and it was not until 1959, at age 25, that he won his first British titles – the RWA's 10 miles road title and the AAA's 2 miles and 7 miles track titles. 1 From then on, he was pretty much unbeatable in England, but the British race most people remember is, interestingly, a loss rather than a victory. In spectacle, excitement and sheer athleticism, the 1960 AAA 2 mile duel between Stan Vickers and Ken stands comparison with any of the great races in the history of the championships. -

The ARTS Bicycle and Pedestrian Plan Update
“The ARTS Bicycle and Pedestrian Plan Update envisions a seamless network of safe and inviting bicycling and walking paths, trails, and on-street facilities, between South Carolina, Georgia and the four member counties, that equitably supports economic development, active transportation, healthy lifestyles and improved quality of life for all citizens and visitors of the region.” Chapter V Two ision , Goa ls, and Objectives 1.1. Objective: Ensure that accommodations for Introduction bicyclists and pedestrians are provided on Based on goals and objectives of existing local all appropriate infrastructure projects where and regional planning documents, the input of pedestrians and bicyclists are permitted to the Project’s steering committee, the project travel. purpose, and relevant examples from around 1.2. Objective: Integrate bicycle and pedes- the country, vision, goals, and objectives are trian facilities in their projects, including, but listed below. The goals and objectives are not limited to, transit, development, public categorized by five of the six E’s associated works, infrastructure, and recreation facili- with bicycle- and walk-friendly community ties. designations. The five E’s are: Engineering, 1.3. Objective: Improve the level of service for Education, Encouragement, Enforcement, and existing bicycle and pedestrian facilities in Evaluation. Equity is considered a sixth E and the member counties. is interwoven within the goals and objectives 1.4. Objective: Increase the mileage of bicycle provided. Objectives 1.6, 1.7, and 3.3 give and pedestrian facilities by fifteen percent particular attention to equity, though it should in each of the region’s four counties within be addressed within the implementation of the next 5 years. -

Energy and Training Module ITU Competitive Coach
37 energy and training module ITU Competitive Coach Produced by the International Triathlon Union, 2007 38 39 energy & training Have you ever wondered why some athletes shoot off the start line while others take a moment to react? Have you every experienced a “burning” sensation in your muscles on the bike? Have athletes ever claimed they could ‘keep going forever!’? All of these situations involve the use of energy in the body. Any activity the body performs requires work and work requires energy. A molecule called ATP (adenosine triphosphate) is the “energy currency” of the body. ATP powers most cellular processes that require energy including muscle contraction required for sport performance. Where does ATP come from and how is it used? ATP is produced by the breakdown of fuel molecules—carbohydrates, fats, and proteins. During physical activity, three different processes work to split ATP molecules, which release energy for muscles to use in contraction, force production, and ultimately sport performance. These processes, or “energy systems”, act as pathways for the production of energy in sport. The intensity and duration of physical activity determines which pathway acts as the dominant fuel source. Immediate energy system Fuel sources ATP Sport E.g. carbohydrates, energy performance proteins, fats “currency” Short term energy system E.g. swimming, cycling, running, transitions Long term energy system During what parts of a triathlon might athletes use powerful, short, bursts of speed? 1 2 What duration, intensity, and type of activities in a triathlon cause muscles to “burn”? When in a triathlon do athletes have to perform an action repeatedly for longer than 10 or 15 3 minutes at a moderate pace? 40 energy systems Long Term (Aerobic) System The long term system produces energy through aerobic (with oxygen) pathways. -

Temporary Traffic Control Zone Pedestrian Access Considerations
Guidance Sheet - Temporary Traffic Control Zone Maintaining Pedestrian Pathways in TTC Zones If a project allows pedestrian access through part of the TTC zone, the pathway should be properly Pedestrian Access Considerations maintained. Note that tape, rope, or a plastic chain strung between devices is not detectable; their use does not comply with the design standards in the ADA or the MUTCD, and these items should not be used as a control for pedestrian movements. When implemented, the following recommendations should improve When developing temporary traffic control (TTC) plans, the importance of pedestrian access in and around safety and convenience: the work zone is often overlooked or underestimated. A basic requirement of work zone traffic control, as provided in the Manual on Uniform Traffic Control Devices (MUTCD), is that the needs of pedestrians, v Inspect pathways regularly, and keep them clear of debris and well-maintained. including those with disabilities, must be addressed in the TTC process. Pedestrians should be provided with reasonably safe, convenient, and accessible paths that replicate as nearly as practical the most v Treat surfaces with non-slip materials for inclement weather. desirable characteristics of the existing sidewalks or footpaths. It is essential to recognize that pedestrians are reluctant to retrace their steps to a prior intersection for a crossing, or to add distance or out-of-the-way v Replace walkway surfaces with holes, cracks, or vertical separation. travel to a destination. This guidance sheet serves to remind TTC designers and construction personnel of v Inspect detour pathways regularly for adequacy of signal timing, signs, and potential traffic the importance of pedestrian access, to stress the need for pedestrian safety, and to offer suggestions that will improve the visibility of pedestrian access. -

Events and Technical Specifications for Athletics
EVENTS AND TECHNICAL SPECIFICATIONS FOR ATHLETICS Track & Field Multi / Combined Events Cross Country Running Road Running Race Walking (see Track & Field) MEN AND WOMEN AGE GROUPS Junior Development – Boys 12-13 yrs (Bantam) – Javelin weight change Midget - ages 14 & 15 Youth - ages 16 & 17 Junior - ages 18 & 19 Senior - ages 20 to 34 for Track & Field Senior – ages 20 to 34 for Out of Stadia Road, Cross Country, Mountain & Trail Running Masters Track & Field – ages 35 & over Masters Road and Cross Country Running - ages 35 & over Note: Event Changes and Additions are highlighted Updated May 2013 BC Athletics is support by BC ATHLETICS TRACK & FIELD CHAMPIONSHIP EVENTS BY AGE CLASS Track & Field Events contested in each age group - 9 years to Masters (35 & over for T&F and 40 & over for Road and Cross Country Running). Unless noted, events are for Females and Males. Ages are as of Dec 31 in the year of competition. Note: Masters Ages are based on the individual’s age on the day or in multiple day events, the first day of competition. Changes / Additions / Clarification: (highlighted) See the specific events and age groups for details Confirmed Changes as of January 1, 2013: Masters ages for Out of Stadia and In-Stadia Events is 35+ 14 & 15 Year Olds in All BC Athletics Competitions inclusive of Championships and Except for BC Summer Games, shall compete as separate age groups and be awarded separately. i.e. Midget 14 events and Midget 15 events. 14 & 15 Year Olds in the BC Summer Games and the BC Summer Games Trials shall compete together as one age group and be awarded as one age group. -

Pedestrian Crossings: Uncontrolled Locations
Pedestrian Crossings: Uncontrolled Locations CENTER FOR TRANSPORTATION STUDIES Pedestrian Crossings: Uncontrolled Locations June 2014 Published By Minnesota Local Road Research Board (LRRB) Web: www.lrrb.org MnDOT Office of Maintenance MnDOT Research Services Section MS 330, 395 John Ireland Blvd. St. Paul, Minnesota 55155 Phone: 651-366-3780 Fax: 651-366-3789 E-mail: [email protected] Acknowledgements The financial and logistical support provided by the Minnesota Local DATA COLLECTION Road Research Board, the Minnesota Department of Transportation (MnDOT), and the Minnesota Local Technical Assistance Program John Hourdos and Stephen Zitzow, University of Minnesota (LTAP) at the Center for Transportation Studies (CTS), University of PRODUCTION Minnesota for this work is greatly acknowledged. Research, Development, and Writing: Bryan Nemeth, Ross Tillman, The procedures presented in this report were developed based on infor- Jeremy Melquist, and Ashley Hudson, Bolton & Menk, Inc. mation from previously published research studies and reports and newly collected field data. Editing: Christine Anderson, CTS The authors would also like to thank the following individuals and orga- Graphic Design: Abbey Kleinert and Cadie Wright Adikhary, CTS, and nizations for their contributions to this document. David Breiter, Bolton & Menk, Inc. TECHNICAL ADVISORY PANEL MEMBERS Tony Winiecki , Scott County Pete Lemke, Hennepin County Kate Miner, Carver County Tim Plath, City of Eagan Mitch Rasmussen, Scott County Jason Pieper, Hennepin County Mitch Bartelt, MnDOT This material was developed by Bolton & Menk, Inc., in coordination with the Minne- Melissa Barnes, MnDOT sota Local Road Research Board for use by practitioners. Under no circumstances shall Tim Mitchell, MnDOT this guidebook be sold by third parties for profit. -

Shoes Approved by World Athletics - As at 01 October 2021
Shoes Approved by World Athletics - as at 01 October 2021 1. This list is primarily a list concerns shoes that which have been assessed by World Athletics to date. 2. The assessment and whether a shoe is approved or not is determined by several different factors as set out in Technical Rule 5. 3. The list is not a complete list of every shoe that has ever been worn by an athlete. If a shoe is not on the list, it can be because a manufacturer has failed to submit the shoe, it has not been approved or is an old model / shoe. Any shoe from before 1 January 2016 is deemed to meet the technical requirements of Technical Rule 5 and does not need to be approved unless requested This deemed approval does not prejudice the rights of World Athletics or Referees set out in the Rules and Regulations. 4. Any shoe in the list highlighted in blue is a development shoe to be worn only by specific athletes at specific competitions within the period stated. NON-SPIKE SHOES Shoe Company Model Track up to 800m* Track from 800m HJ, PV, LJ, SP, DT, HT, JT TJ Road* Cross-C Development Shoe *not including 800m *incl. track RW start date end date ≤ 20mm ≤ 25mm ≤ 20mm ≤ 25mm ≤ 40mm ≤ 25mm 361 Degrees Flame NO NO NO NOYES NO Adidas Adizero Adios 3 NO YES NO YES YES YES Adidas Adizero Adios 4 NO YES NO YES YES YES Adidas Adizero Adios 5 NO YES NO YES YES YES Adidas Adizero Adios 6 NO YES NO YES YES YES Adidas Adizero Adios Pro NO NO NO NOYES NO Adidas Adizero Adios Pro 2 NO NO NO NOYES NO Adidas Adizero Boston 8 NO NO NO NOYES NO Adidas Adizero Boston 9 NO NO NO -

1 MEASURING ROAD RUNNING COURSES by Ted Corbitt New York
MEASURING ROAD RUNNING COURSES by Ted Corbitt New York Pioneer Club Contributors Jack Barry, Merchantville, N. J. B. Bright, Indianapolis, Indiana Robert Campbell, W. Roxbury, Mass. P. W. Cerutty, Portsea, Victoria, Australia Neil Farrell, Hamilton, Ontario, Canada Jack Girling, Hamilton, Ontario, Canada Scott Hamilton, Anchorage, Alaska Philip Jachelski, Baltimore, Maryland Hugh Jascourt, Arlington, Va. John Jewell, Wokingham, Berks., England Gordon McKenzie, Great Neck, Long Island, New York A. L. Monteverde, Los Angeles, California Stan Pisani, Newtown, N. S. W., Australia Bob Prentice, Melbourne, Australia C. A. Robbins, M. D., Middletown, Connecticut W. M. Ruthrauff, D. Sc., Philadelphia, Pa. John Sterner, Bronx, N. Y. S. Takenaka, Tokyo, Japan Geoff Watt, Melbourne, Australia Bill Wiklund, Clifton, N. J. The author acknowledges and appreciates the special assistance given in this project by John Sterner, Bronx, N. Y. and by John Jewell, Road Runners Club of England. * * * * * PROVERBS 20 #10 - "Divers weights, and divers measures, both of them are alike an abomination to the Lord." * * * * * ROAD RUNNERS CLUB, USA Publication No. 4 August 1964 Price $1.00 Any profits from the sale of this monograph will be used to promote accurate measurements of road running courses in the USA. 1 CONTENTS SUBJECT PAGE Introduction 3 Road Running Courses 4 Methods of Measuring Road Racing Courses 6 Where to Measure a Road Course 6 Accuracy 7-8 Measure Conversions 9 Standard Distance 9 Commonly Used Methods of Measuring Distances for Road Race Courses 10 Recommended Methods of Measuring Road Courses 10 I. Surveying 10 II. Tape Measurements 11-17 III. Calibrated Cycle Method 18-21 IV. -

Long Distance Running Division
2006 Year-End Reports 28th Annual Meeting Reports from the Long Distance Running Division Men’s Long Distance Running Women’s Long Distance Running Masters Long Distance Running Cross Country Council Mountain, Ultra & Trail (MUT) Council Road Running Technical Council 97 National Officers, National Office Staff, Division and Committee Chairs 98 2006 Year-End Reports 28th Annual Meeting Men’s Long Distance Running B. USA National Championships 2005 USA Men's 10 km Championship – Food KEY POINTS World Senior Bowl 10k Mobile, AL – November 5, 2005 Update October 2005 to December 2005 http://www.usatf.org/events/2005/USA10kmCha As last year’s USATF Men’s LDR Annual Report mpionship-Men/ was written in October 2005 in order to meet A dominant display and new course record of publication deadlines for the Annual Convention, 28:11 for Dathan Ritzenhein to become the USA here are a few highlights of Men’s activities from National Champion. October 2005 through to the end of 2005. (Web site links provided where possible.) 2005 USATF National Club Cross Country Championships A. Team USA Events November 19, 2005 Genesee Valley Park - IAAF World Half Marathon Championships – Rochester, NY October 1, 2005, Edmonton, Canada http://www.usatf.org/events/2005/USATFClubX http://www.usatf.org/events/2005/IAAFWorldHalf CChampionships/ MarathonChampionships/ An individual win for Matt Tegenkamp, and Team Scores of 1st Hansons-Brooks D P 50 points th 6 place team United States - 3:11:38 - 2nd Asics Aggie R C 68 points USA Team Leader: Allan Steinfeld 3rd Team XO 121 points th 15 Ryan Shay 1:03:13 th 20 Jason Hartmann 1:03:32 C. -

Pacing in Time-Limited Ultramarathons from 6 to 24 Hours—The Aspects of Age, Sex and Performance Level
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2021 Pacing in Time-Limited Ultramarathons from 6 to 24 Hours—The Aspects of Age, Sex and Performance Level Deusch, Hagen ; Nikolaidis, Pantelis T ; Alvero-Cruz, José Ramón ; Rosemann, Thomas ; Knechtle, Beat Abstract: (1) Background: Compared with marathon races, pacing in time-limited ultramarathons has only been poorly discussed in the literature. The aim of the present study was to analyze the interaction of performance level, age and sex with pacing during 6 h, 12 h or 24 h time-limited ultramarathons. (2) Methods: Participants (n = 937, age 48.62 ± 11.80 years) were the finishers in 6 h (n = 40, 17 women and 23 men), 12 h (n = 232, 77 women and 155 men) and 24 h (n = 665, 166 women and 409 men) ultramarathons. The coefficient of variation (CV), calculated as SD/mean, was used to described pacing. Low scores of CV denoted a more even pacing, and vice versa. A two-way analysis of variance examined the main effects and interactions of sex and race duration on age, race speed and pacing. (3) Results: More men participated in the longer race distances than in the shorter ones and men were older and faster than women. Comparing the 6 h, 12 h and 24 h races, the finishers in the 6 h were the fastest, the finishers in the 12 h were the oldest and the finishers in the 24 h showed the most variablepacing. Furthermore, the faster running speed in the 12 h (women, r = −0.64; men, r = −0.49, p < 0.001) and the 24 h (r = −0.47 in women and men, p < 0.001) was related to less variable pacing. -
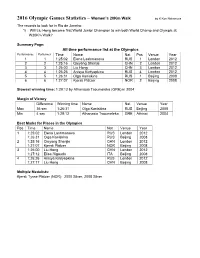
2016 Olympic Games Statistics
2016 Olympic Games Statistics – Women’s 20Km Walk by K Ken Nakamura The records to look for in Rio de Janeiro: 1) Will Liu Hong become first World Junior Champion to win both World Champ and Olympic at W20Km Walk? Summary Page: All time performance list at the Olympics Performance Performer Time Name Nat Pos Venue Year 1 1 1:25:02 Elena Lashmanova RUS 1 London 2012 2 2 1:25:16 Qieyang Shenjie CHN 2 London 2012 3 3 1:26:00 Liu Hong CHN 3 London 2012 4 4 1:26:26 Anisya Kirdyapkina RUS 4 London 2012 5 5 1:26:31 Olga Kaniskina RUS 1 Beijing 2008 6 6 1:27:07 Kjersti Plätzer NOR 2 Beijing 2008 Slowest winning time: 1:29:12 by Athanasia Tsoumeleka (GRE) in 2004 Margin of Victory Difference Winning time Name Nat Venue Year Max 36 sec 1:26:31 Olga Kaniskina RUS Beijing 2008 Min 4 sec 1:29:12 Athanasia Tsoumeleka GRE Athinai 2004 Best Marks for Places in the Olympics Pos Time Name Nat Venue Year 1 1:25:02 Elena Lashmanova RUS London 2012 1:26:31 Olga Kaniskina RUS Beijing 2008 2 1:25:16 Qieyang Shenjie CHN London 2012 1:27:07 Kjersti Plätzer NOR Beijing 2008 3 1:26:00 Liu Hong CHN London 2012 1:27:12 Elisa Rigaudo ITA Beijing 2008 4 1:26:26 Anisya Kirdyapkina RUS London 2012 1:27:17 Liu Hong CHN Beijing 2008 Multiple Medalists: Kjersti Tysse Plätzer (NOR): 2000 Silver, 2008 Silver All time performance list at the Olympics Performance Performer Time Name Nat Pos Venue Year 1 1 1:25:02 Elena Lashmanova RUS 1 London 2012 2 2 1:25:16 Qieyang Shenjie CHN 2 London 2012 3 3 1:26:00 Liu Hong CHN 3 London 2012 4 4 1:26:26 Anisya Kirdyapkina RUS 4 London -

The International Ski Competition Rules (Icr)
THE INTERNATIONAL SKI COMPETITION RULES (ICR) BOOK II CROSS-COUNTRY APPROVED BY THE 51ST INTERNATIONAL SKI CONGRESS, COSTA NAVARINO (GRE) EDITION MAY 2018 INTERNATIONAL SKI FEDERATION FEDERATION INTERNATIONALE DE SKI INTERNATIONALER SKI VERBAND Blochstrasse 2; CH- 3653 Oberhofen / Thunersee; Switzerland Telephone: +41 (33) 244 61 61 Fax: +41 (33) 244 61 71 Website: www.fis-ski.com ________________________________________________________________________ All rights reserved. Copyright: International Ski Federation FIS, Oberhofen, Switzerland, 2018. Oberhofen, May 2018 Table of Contents 1st Section 200 Joint Regulations for all Competitions ................................................... 3 201 Classification and Types of Competitions ................................................... 3 202 FIS Calendar .............................................................................................. 5 203 Licence to participate in FIS Races (FIS Licence) ...................................... 7 204 Qualification of Competitors ....................................................................... 8 205 Competitors Obligations and Rights ........................................................... 9 206 Advertising and Sponsorship .................................................................... 10 207 Competition Equipment and Commercial Markings .................................. 12 208 Exploitation of Electronic Media Rights .................................................... 13 209 Film Rights ..............................................................................................