Multi-Scale Studies of Populations, Distribution and Habitat Associations of Marbled Murrelets in Clayoquot Sound, British Columbia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

KEEP IT LOCAL Spring & Summer Getaways Close to Home a WHALE
ExperiencesESCORTED GROUP TRAVEL SINCE 1972 Vol 09 – Spring 2021 KEEP IT LOCAL A WHALE OF A TIME PLAN NOW, GO LATER spring & summer getaways whale watching exciting new close to home adventures in Canada global itineraries THE WELLS GRAY TOURS ADVANTAGE WE PLAN | YOU PACK | NO WORRIES escorted tours with local offices with early booking discounts ExperienceAWE-INSPIRING ADVENTURES AHEAD experienced tour directors friendly, helpful staff and loyalty rewards program Spring is here and thoughts turn to travel. I think a great It seems so routine to be doing Zoom meetings and, LS GRAY EL T poet said that or maybe I have paraphrased. In any case, even though it is tiring looking at a screen for so many W O I hope this is true for you. Our phones are staying busy hours, it saves a lot of time travelling to in-person U pick-up points in BC Interior, single, double and triple R and lots of bookings are coming in, especially now that meetings. One day recently, I had four Zoom meetings P h S Vancouver Island, and fares available R vaccinations are starting earlier for most customers. We including my yoga class which for safety has moved from O b a i Lower Mainland V d are anxiously awaiting the lifting of travel restrictions by a studio to online. I n D i p I y N Dr. Henry, so our British Columbia tours can hopefully i s Although I am working from home every day, I wonder G m c 2 proceed starting in late April. -

An Examination of Nuu-Chah-Nulth Culture History
SINCE KWATYAT LIVED ON EARTH: AN EXAMINATION OF NUU-CHAH-NULTH CULTURE HISTORY Alan D. McMillan B.A., University of Saskatchewan M.A., University of British Columbia THESIS SUBMI'ITED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in the Department of Archaeology O Alan D. McMillan SIMON FRASER UNIVERSITY January 1996 All rights reserved. This work may not be reproduced in whole or in part, by photocopy or other means, without permission of the author. APPROVAL Name: Alan D. McMillan Degree Doctor of Philosophy Title of Thesis Since Kwatyat Lived on Earth: An Examination of Nuu-chah-nulth Culture History Examining Committe: Chair: J. Nance Roy L. Carlson Senior Supervisor Philip M. Hobler David V. Burley Internal External Examiner Madonna L. Moss Department of Anthropology, University of Oregon External Examiner Date Approved: krb,,,) 1s lwb PARTIAL COPYRIGHT LICENSE I hereby grant to Simon Fraser University the right to lend my thesis, project or extended essay (the title of which is shown below) to users of the Simon Fraser University Library, and to make partial or single copies only for such users or in response to a request from the library of any other university, or other educational institution, on its own behalf or for one of its users. I further agree that permission for multiple copying of this work for scholarly purposes may be granted by me or the Dean of Graduate Studies. It is understood that copying or publication of this work for financial gain shall not be allowed without my written permission. -

Strathcona Park Trails Guide Special
STRATHCONA PARK TRAILS GUIDE SPECIAL Hiking Guide NootkaWILDWILD Island ISLEISLE Canada’sCanada’s WestWest CoastCoast AdventureAdventure MagazineMagazine ININ THETHE WAKEWAKE OFOF CAPTAINCAPTAIN VANCOUVERVANCOUVER A CLIMBER’S LIFELIFE CAVINGCAVING ININ THAILANDTHAILAND 3Chancesto WIN Apairoftickets to FREE ISSUE #12 AUGUST-SEPTEMBER, 2000 see page 26 for details www.wildislemagazine.bc.ca HIKE NOOTKA ISLAND The Resort at Cape Mudge Quadra Island BEACHFRONT CABIN . Only Getaway .95* September 16 - 30, 2000 $119 * based on quad occupancy 2 Bedroom With Jacuzzi Bathtub Gas Fireplace Kitchenette Us! 1-800-665-7745 [email protected] Tel: (250) 285-2042 Fax: (250) 285-2532 CCONTENTONTENTSS Cover: Curtis Lyon, South Ridge of Victoria Peak. Photo: Philip Stone BC Parks View Point p9 Volunteering in Paradise Gabe Jutras, Kwisatz Haderach 5.11d, Crest Creek Photo: Josie Boulding Linvin’ the Dream p10 Contents: The life of the Surfer at Long Beach. unsponsored climber Photo: Corrie Wright Strathcona Park p13 An overview of the hiking trails in BC’s oldest park Thai Caves p17 Islanders Abroad Underground adventure in Thailand Mt Colonel Foster p19 Guide to Vancouver Island’s alpine climbing mecca Benighted on Mt Klitsa p21 A lesson earned and a lesson learned In the Wake of Vancouver p22 The explorations of Cpt Vancouver on BC’s west coast. Happenings p26 Coming events on the west coast 3Chancesto WIN Apairoftickets to see page 26 for details west coast adventure magazine • www.wildislemagazine.bc.ca • CENTRE PULLOUT - NOOTKA ISLAND - HIKING -

British Columbia Regional Guide Cat
National Marine Weather Guide British Columbia Regional Guide Cat. No. En56-240/3-2015E-PDF 978-1-100-25953-6 Terms of Usage Information contained in this publication or product may be reproduced, in part or in whole, and by any means, for personal or public non-commercial purposes, without charge or further permission, unless otherwise specified. You are asked to: • Exercise due diligence in ensuring the accuracy of the materials reproduced; • Indicate both the complete title of the materials reproduced, as well as the author organization; and • Indicate that the reproduction is a copy of an official work that is published by the Government of Canada and that the reproduction has not been produced in affiliation with or with the endorsement of the Government of Canada. Commercial reproduction and distribution is prohibited except with written permission from the author. For more information, please contact Environment Canada’s Inquiry Centre at 1-800-668-6767 (in Canada only) or 819-997-2800 or email to [email protected]. Disclaimer: Her Majesty is not responsible for the accuracy or completeness of the information contained in the reproduced material. Her Majesty shall at all times be indemnified and held harmless against any and all claims whatsoever arising out of negligence or other fault in the use of the information contained in this publication or product. Photo credits Cover Left: Chris Gibbons Cover Center: Chris Gibbons Cover Right: Ed Goski Page I: Ed Goski Page II: top left - Chris Gibbons, top right - Matt MacDonald, bottom - André Besson Page VI: Chris Gibbons Page 1: Chris Gibbons Page 5: Lisa West Page 8: Matt MacDonald Page 13: André Besson Page 15: Chris Gibbons Page 42: Lisa West Page 49: Chris Gibbons Page 119: Lisa West Page 138: Matt MacDonald Page 142: Matt MacDonald Acknowledgments Without the works of Owen Lange, this chapter would not have been possible. -

The Fur Trade and Early Capitalist Development in British Columbia
THE FUR TRADE AND EARLY CAPITALIST DEVELOPMENT IN BRITISH COLUMBIA RENNIE WARBURTON, Department of Sociology, University of Victoria, Victoria, British Columbia, Canada, V8W 2Y2. and STEPHEN SCOTT, Department of Sociology and Anthropology, Carleton University, Ottawa, Ontario, Canada, K1S 5B6. ABSTRACT/RESUME Although characterized by unequal exchange, the impact of the fur trade on the aboriginal societies of what became British Columbia involved minimal dis- ruption because the indigenous modes of production were easily articulated with mercantile capitalism. It was the problems arising from competition and increasing costs of transportation that led the Hudson's Bay Company to begin commodity production in agriculture, fishing and lumbering, thereby initiating capitalist wage-labour relations and paving the way for the subsequent disast- rous decline in the well-being of Native peoples in the province. Bien que characterisé par un échange inégale, l'impact du commerce de fourrure sur les societiés aborigonaux sur ce qui est devenu la Colombie Britanique ne dérangèrent pas les societés, car les modes indigènes de production était facile- ment articulés avec un capitalisme mercantile. Ce sont les problèmes qui venaient de la competition et les frais de transportation qui augmentaient qui mena la Companie de la Baie d'Hudson à commencer la production de commodités dans les domaines de l'agriculture, la pêche et l'exploitement du bois. Par ce moyen elle initia des rapports de salaire-travail capitalist et prépara les voies pour aboutir å une reduction catastrophique du bien-être des natifs dans cette province. THE CANADIAN JOURNAL OF NATIVE STUDIES V, 1(1985):27-46 28 R E N N I E W A R B U R T O N / S T E P H E N S C O T T INTRODUCTION In the diverse cultures in British Columbia prior to and after contact with the Europeans, economic activity included subsistence hunting, fishing and gathering as well as domestic handicrafts. -

Sailing Directions (Enroute)
PUB. 154 SAILING DIRECTIONS (ENROUTE) ★ BRITISH COLUMBIA ★ Prepared and published by the NATIONAL GEOSPATIAL-INTELLIGENCE AGENCY Bethesda, Maryland © COPYRIGHT 2007 BY THE UNITED STATES GOVERNMENT NO COPYRIGHT CLAIMED UNDER TITLE 17 U.S.C. 2007 TENTH EDITION For sale by the Superintendent of Documents, U.S. Government Printing Office Internet: http://bookstore.gpo.gov Phone: toll free (866) 512-1800; DC area (202) 512-1800 Fax: (202) 512-2250 Mail Stop: SSOP, Washington, DC 20402-0001 Preface 0.0 Pub. 154, Sailing Directions (Enroute) British Columbia, 0.0NGA Maritime Domain Website Tenth Edition, 2007, is issued for use in conjunction with Pub. http://www.nga.mil/portal/site/maritime 120, Sailing Directions (Planning Guide) Pacific Ocean and 0.0 Southeast Asia. Companion volumes are Pubs. 153, 155, 157, 0.0 Courses.—Courses are true, and are expressed in the same 158, and 159. manner as bearings. The directives “steer” and “make good” a 0.0 Digital Nautical Chart 26 provides electronic chart coverage course mean, without exception, to proceed from a point of for the area covered by this publication. origin along a track having the identical meridianal angle as the 0.0 This publication has been corrected to 21 July 2007, includ- designated course. Vessels following the directives must allow ing Notice to Mariners No. 29 of 2007. for every influence tending to cause deviation from such track, and navigate so that the designated course is continuously Explanatory Remarks being made good. 0.0 Currents.—Current directions are the true directions toward 0.0 Sailing Directions are published by the National Geospatial- which currents set. -

Clayoquot Sound)
.. Catalogue of Salmon Streams and Spawning Escapements of Statistical Area 24 ( Clayoquot Sound) R.F Brown, M.J. Comfort, & D.E. Marshall . Fisheries &Oceans Enhancement Services Branch 1090 West Pender St. Vancouver. B. C. V6E 2P1 December 1979 Fisheries & Marine Service Data Report No. 80 Fisheries and Marine Service Data Reports These reports provide a medium for filing and archiving data compilations where little or no analysis is included. Such compilations commonly will have been prepared in support of other journal publications or reports. The subject matter of Data Reports reflects the broad interests and policies of the Fisheries and M arine Service, namely, fisheries management, technology and development, ocean sciences, and aquatic environments relevant to Canada. Numbers 1-25 in this series were issued as Fisheries and Marine Service Data Records by the Pacific Biological Station, N anaimo, B.C The series name was changed with report number 26. Data Reports are not intended for general distribution and the contents must not be referred to in other publications without prior written clearance from the Issuing establishment. The correct citation appears above the abstract of each report. Service des peches et de la mer Rapports statistiques Ces rapports servent de base a la compilation des donnees de classel11ent et d'archives pour lesquelles iI y a peu ou point d'analyse. Celte compilation aura d'ordinaire ete preparee pour appuyer d'autres publications ou rapports. Les sujets des Rapports statistiques refietent la vaste gamme des interets et politiques du Service des peches et de la mer, notamment gestion des peches, techniques et developpement, sciences oceaniques et environnements aquatiques, au Canada . -

You Are in Wolf and Cougar Country
t Keep Predators Wild & Wary ... Stay Safe! Parks Canada needs YOUR help to If you encounter a wolf or cougar: prevent people-predator conflicts and Are People at Risk? • Pick up small children. to keep our predators wild! Cougars very rarely prey on people. Children and crouching adults are more at • Gather the group together. risk of attack as they more closely resemble prey. People on their own are also • Do not run. Wolves and cougars are native to Vancouver Island, more at risk than groups of people. Wolf attacks are even rarer. • Do not crouch down. and as predators, are vital to a healthy coastal In this region, there have been three cougar attacks. In each case children were • Make and maintain eye contact. ecosystem. They may be encountered anywhere in attacked: one on the West Coast Trail in 1985, one fatal attack just outside the • Wave your arms and shout. Pacific Rim National Park Reserve. National parks are park in Clayoquot Sound in 1989 and another at Kennedy Lake in 2011. • Do all you can to appear larger and to scare the great places to view wildlife in their natural habitat. animal away. However, once animals become accustomed to In 2000, a wolf that had obtained food from previous campers, attacked a © Parks Canada / Josh McCulloch 2009 • Avoid scaring the animal into the path of other © Parks Canada / Josh McCulloch 2004 sleeping man by a campfire in Clayoquot Sound. people they are in danger of losing their wildness. Keep kids close. people. The repeated presence of humans that brings no Keep kayak hatches secure. -

Coastal Waterbird Population Trends in the Strait of Georgia 1999–2011: Results from the First 12 Years of the British Columbia Coastal Waterbird Survey
8 Coastal waterbird trends - Crewe et al. Coastal waterbird population trends in the Strait of Georgia 1999–2011: Results from the first 12 years of the British Columbia Coastal Waterbird Survey Tara Crewe1, Karen Barry2, Pete Davidson2,3, Denis Lepage1 1 Bird Studies Canada - National Headquarters, PO Box. 160, Port Rowan, Ont. N0E 1M0 2 Bird Studies Canada – British Columbia Program, 5421 Robertson Road, RR1, Delta, B.C. V4K 3N2; e-mail: [email protected] 3 Corresponding author Abstract: The British Columbia Coastal Waterbird Survey is a citizen science long-term monitoring program implemented by Bird Studies Canada to assess population trends and ecological needs of waterbirds using the province’s coastal and inshore marine habitats. Standard monthly counts from more than 200 defined sites within the Strait of Georgia were analysed using route-regression techniques to estimate population indices and assess trends in 57 waterbird species over a 12-year period spanning the non-breeding periods from 1999–2000 to 2010–11. A power analysis was also conducted to validate the rigor of the survey design. Results indicate that the survey is detecting annual changes of 3% or less for populations of 29 waterbirds of a wide variety of guilds. Thirty-three species showed stable populations or no trend, 22 species showed significantly declining trends, and just three species showed significant increasing trends. We evaluate these results in the context of other long-term monitoring initiatives in the Salish Sea, highlighting specific birds to watch from a conservation perspective. Among those that showed a declining trend were a guild of piscivores, including Western and Horned Grebes, Common, Red-throated and Pacific Loons, and Rhinoceros Auklet; several sea ducks (Black and White-winged Scoters, Long-tailed Duck, Barrow’s Goldeneye, Harlequin Duck); two shorebirds (Dunlin, Surfbird); and Great Blue Heron. -

Vital Signs Report
CLAYOQUOT SOUND BIOSPHERE REGION’S 2018 Welcome to the Clayoquot Sound Biosphere Region’s Vital Signs® 2018 Table of Contents From the Vital Signs Research Team About Vital Signs 2 “We hope the 2018 Vital Signs report ¸ Grounded in the Nuu-chah-nulth (nuucaanuł) ¸ ˇ informs and inspires dialogue and principle of hišukniš cawaak, everything is one, Vital Our Region 3 collaboration to further our collective Signs 2018 can help us to understand the complex Cycle of Poverty efforts to build healthy communities and changing systems in which we live and the necessary pathways we need to navigate in order to in Our Region: and achieve sustainable development.” Inspiring Action support sustainable ecosystems and¸ communities. One of these pathways is nuucaanułˇ language for Change 4 Tammy Dorward and Catherine Thicke revitalization. This year, we’ve worked with a regional Co-chairs, Board of Directors Environment 5-6 committee of elders¸ and language keepers to incor- Clayoquot Biosphere Trust porate nuucaanułˇ throughout the report. Climate Change Impacts 7-8 We’ve collected a range of local data to highlight pri- ority areas for community-wide action and listened People & Work 9 From our Executive Director closely to community concerns. We’ve heard that Income Inequality 10 I am pleased to present our 2018 Vital Signs report. our young people are struggling with mental health Vital Signs is a valuable tool for understanding our issues and that they lack youth programs. Families Housing 11 progress toward achieving all aspects of sustainabili- are challenged with rising housing costs and the ty—cultural, social, economic, and environmental. -

PROVINCI L Li L MUSEUM
PROVINCE OF BRITISH COLUMBIA REPORT OF THE PROVINCI_l_Li_L MUSEUM OF NATURAL HISTORY • FOR THE YEAR 1930 PRINTED BY AUTHORITY OF THE LEGISLATIVE ASSEMBLY. VICTORIA, B.C. : Printed by CHARLES F. BANFIELD, Printer to tbe King's Most Excellent Majesty. 1931. \ . To His Honour JAMES ALEXANDER MACDONALD, Administrator of the Province of British Columbia. MAY IT PLEASE YOUR HONOUR: The undersigned respectfully submits herewith the Annual Report of the Provincial Museum of Natural History for the year 1930. SAMUEL LYNESS HOWE, Pt·ovincial Secretary. Pt·ovincial Secretary's Office, Victoria, B.O., March 26th, 1931. PROVINCIAl. MUSEUM OF NATURAl. HISTORY, VICTORIA, B.C., March 26th, 1931. The Ho1Wm·able S. L. Ho11ie, ProvinciaZ Secreta11}, Victo1·ia, B.a. Sm,-I have the honour, as Director of the Provincial Museum of Natural History, to lay before you the Report for the year ended December 31st, 1930, covering the activities of the Museum. I have the honour to be, Sir, Your obedient servant, FRANCIS KERMODE, Director. TABLE OF CONTENTS . PAGE. Staff of the Museum ............................. ------------ --- ------------------------- ----------------------------------------------------- -------------- 6 Object.. .......... ------------------------------------------------ ----------------------------------------- -- ---------- -- ------------------------ ----- ------------------- 7 Admission .... ------------------------------------------------------ ------------------ -------------------------------------------------------------------------------- -
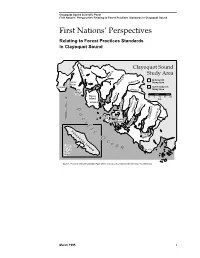
First Nations' Perspectives
Clayoquot Sound Scientific Panel Trigger Type Type First Nations’ Perspectives Relating to Forest Practices Standards in Clayoquot Sound First Nations’ Perspectives Relating to Forest Practices Standards in Clayoquot Sound Sydney R. Clayoquot Sound Study Area Megin R. included in Hesquiat Bedwell R. Study Area Harbour Moyeha R. Hesquiat not included in Hot Ursus R. Study Area Springs Cove Flores 0 5 10 15 20 N Island km Cypre R. Ahousat P Bulson Cr. Herbert Inlet Tofino Cr. a Tranquil Cr. Vargas Opitsat c Island Clayoquot R. Meares Kennedy R. i Tofino Island f Tofino Inlet i Vancouver Islandc Kennedy O Lake c e Clayoquot a Sound n Ucluelet Study Area Source: Province of British Columbia (April 1993). Clayoquot Sound Land Use Decision: Key Elements. March 1995 i Clayoquot Sound Scientific Panel Trigger Type Type First Nations’ Perspectives Relating to Forest Practices Standards in Clayoquot Sound March 1995 ii Clayoquot Sound Scientific Panel Trigger Type Type First Nations’ Perspectives Relating to Forest Practices Standards in Clayoquot Sound Table of Contents Acknowledgments ......................................................................................................... v Executive Summary...................................................................................................... vii 1.0 Introduction........................................................................................................... 1 1.1 Context of this Report ...................................................................................