San Joaquin Valley Orcutt Grass)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

Phylogeny and Subfamilial Classification of the Grasses (Poaceae) Author(S): Grass Phylogeny Working Group, Nigel P
Phylogeny and Subfamilial Classification of the Grasses (Poaceae) Author(s): Grass Phylogeny Working Group, Nigel P. Barker, Lynn G. Clark, Jerrold I. Davis, Melvin R. Duvall, Gerald F. Guala, Catherine Hsiao, Elizabeth A. Kellogg, H. Peter Linder Source: Annals of the Missouri Botanical Garden, Vol. 88, No. 3 (Summer, 2001), pp. 373-457 Published by: Missouri Botanical Garden Press Stable URL: http://www.jstor.org/stable/3298585 Accessed: 06/10/2008 11:05 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=mobot. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with the scholarly community to preserve their work and the materials they rely upon, and to build a common research platform that promotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected]. -

Biological Resources Assessment the Ranch ±530- Acre Study Area City of Rancho Cordova, California
Biological Resources Assessment The Ranch ±530- Acre Study Area City of Rancho Cordova, California Prepared for: K. Hovnanian Homes October 13, 2017 Prepared by: © 2017 TABLE OF CONTENTS 1.0 Introduction ......................................................................................................................... 1 1.1. Project Description ........................................................................................................... 1 2.0 Regulatory Framework ........................................................................................................ 2 2.1. Federal Regulations .......................................................................................................... 2 2.1.1. Federal Endangered Species Act ............................................................................... 2 2.1.2. Migratory Bird Treaty Act ......................................................................................... 2 2.1.3. The Bald and Golden Eagle Protection Act ............................................................... 2 2.2. State Jurisdiction .............................................................................................................. 3 2.2.1. California Endangered Species Act ........................................................................... 3 2.2.2. California Department of Fish and Game Codes ...................................................... 3 2.2.3. Native Plant Protection Act ..................................................................................... -

US EPA-Pesticides; Dodine
UNITED STATES ENVIRONMENTAL PROTECTION AGENCY WASHINGTON D.C., 20460 OFFICE OF PREVENTION, PESTICIDESDES AND TOXIC SUBSTANCES PC Code: 044301 DP Barcode: D338148 Date: January 22, 2008 MEMORANDUM SUBJECT: Ecological Risk Assessment for the Dodine Section 3 New Use on Peanuts and Bananas TO: Robert Westin, Product Manager Mary Waller, Team Leader Registration Division (7505P) FROM: Christopher J. Salice, P.h.D, Biologist Marietta Echeverria, Envronmental Scientist Environmental Risk Branch IV Environmental Fate and Effects Division (7507P) REVIEWED BY: Thomas Steeger, Ph.D., Senior Biologist R. David Jones, Ph.D., Senior Agronomist Environmental Risk Branch IV Environmental Fate and Effects Division (7507P) APPROVED BY: Elizabeth Behl, Branch Chief Environmental Risk Branch IV Environmental Fate and Effects Division (7507P) The Environmental Fate and Effects Division (EFED) has reviewed the proposed label for the use of dodine (n-dodecylguanidine monoacetate; CAS 2439-10-3) and its end-use product SYLLIT® FL (39.6% dodine) fungicide on peanuts and bananas. The results of this screening-level risk assessment indicate that the proposed new uses of dodine on peanuts and bananas have the potential for direct adverse effects on listed and non-listed freshwater and estuarine/marine invertebrates, listed and non-listed vascular and non-vascular plants, and listed and non-listed birds and mammals. Major data gaps are listed below. Without these data potential risk to the associated taxa can not be precluded: • Aquatic vascular plant toxicity data (850.4400) There is uncertainty regarding the potential chronic effects of dodine to saltwater invertebrates and fish since there are no toxicity data. Using acute-to-chronic ratios (ACR) from freshwater species to calculate chronic endpoints for the saltwater species, however, suggests that risks may be low. -

Viruses Virus Diseases Poaceae(Gramineae)
Viruses and virus diseases of Poaceae (Gramineae) Viruses The Poaceae are one of the most important plant families in terms of the number of species, worldwide distribution, ecosystems and as ingredients of human and animal food. It is not surprising that they support many parasites including and more than 100 severely pathogenic virus species, of which new ones are being virus diseases regularly described. This book results from the contributions of 150 well-known specialists and presents of for the first time an in-depth look at all the viruses (including the retrotransposons) Poaceae(Gramineae) infesting one plant family. Ta xonomic and agronomic descriptions of the Poaceae are presented, followed by data on molecular and biological characteristics of the viruses and descriptions up to species level. Virus diseases of field grasses (barley, maize, rice, rye, sorghum, sugarcane, triticale and wheats), forage, ornamental, aromatic, wild and lawn Gramineae are largely described and illustrated (32 colour plates). A detailed index Sciences de la vie e) of viruses and taxonomic lists will help readers in their search for information. Foreworded by Marc Van Regenmortel, this book is essential for anyone with an interest in plant pathology especially plant virology, entomology, breeding minea and forecasting. Agronomists will also find this book invaluable. ra The book was coordinated by Hervé Lapierre, previously a researcher at the Institut H. Lapierre, P.-A. Signoret, editors National de la Recherche Agronomique (Versailles-France) and Pierre A. Signoret emeritus eae (G professor and formerly head of the plant pathology department at Ecole Nationale Supérieure ac Agronomique (Montpellier-France). Both have worked from the late 1960’s on virus diseases Po of Poaceae . -

Baja California, Mexico, and a Vegetation Map of Colonet Mesa Alan B
Aliso: A Journal of Systematic and Evolutionary Botany Volume 29 | Issue 1 Article 4 2011 Plants of the Colonet Region, Baja California, Mexico, and a Vegetation Map of Colonet Mesa Alan B. Harper Terra Peninsular, Coronado, California Sula Vanderplank Rancho Santa Ana Botanic Garden, Claremont, California Mark Dodero Recon Environmental Inc., San Diego, California Sergio Mata Terra Peninsular, Coronado, California Jorge Ochoa Long Beach City College, Long Beach, California Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Biodiversity Commons, Botany Commons, and the Ecology and Evolutionary Biology Commons Recommended Citation Harper, Alan B.; Vanderplank, Sula; Dodero, Mark; Mata, Sergio; and Ochoa, Jorge (2011) "Plants of the Colonet Region, Baja California, Mexico, and a Vegetation Map of Colonet Mesa," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 29: Iss. 1, Article 4. Available at: http://scholarship.claremont.edu/aliso/vol29/iss1/4 Aliso, 29(1), pp. 25–42 ’ 2011, Rancho Santa Ana Botanic Garden PLANTS OF THE COLONET REGION, BAJA CALIFORNIA, MEXICO, AND A VEGETATION MAPOF COLONET MESA ALAN B. HARPER,1 SULA VANDERPLANK,2 MARK DODERO,3 SERGIO MATA,1 AND JORGE OCHOA4 1Terra Peninsular, A.C., PMB 189003, Suite 88, Coronado, California 92178, USA ([email protected]); 2Rancho Santa Ana Botanic Garden, 1500 North College Avenue, Claremont, California 91711, USA; 3Recon Environmental Inc., 1927 Fifth Avenue, San Diego, California 92101, USA; 4Long Beach City College, 1305 East Pacific Coast Highway, Long Beach, California 90806, USA ABSTRACT The Colonet region is located at the southern end of the California Floristic Province, in an area known to have the highest plant diversity in Baja California. -

Baseline Biodiversity Report
FINAL Baseline Biodiversity Survey for Potrero Mason Property Prepared for: County of San Diego Department of Parks and Recreation 5500 Overland Avenue Drive, Suite 410 San Diego, California 92123 Contact: Jennifer Price Prepared by: 605 Third Street Encinitas, California 92024 Contact: Brock Ortega DECEMBER 2012 Printed on 30% post-consumer recycled material. Final Baseline Biodiversity Survey Potrero Mason Property TABLE OF CONTENTS Section Page No. LIST OF ACRONYMS ................................................................................................................ V EXECUTIVE SUMMARY .......................................................................................................VII 1.0 INTRODUCTION..............................................................................................................1 1.1 Purpose of the Report.............................................................................................. 1 1.2 MSCP Context ........................................................................................................ 1 2.0 PROPERTY DESCRIPTION ...........................................................................................9 2.1 Project Location ...................................................................................................... 9 2.2 Geographical Setting ............................................................................................... 9 2.3 Geology and Soils .................................................................................................. -

Initial Study with Mitigated Negative Declaration
Merced 140 Roadside Safety Improvements On State Route 140 in Merced County 10-MER-140-2.3/49.0 EA 10-0Y130 and Project ID 1013000243 SCH Number 2019059129 Initial Study with Mitigated Negative Declaration Prepared by the State of California Department of Transportation April 2020 General Information About This Document The California Department of Transportation (Caltrans) prepared this Initial Study with Mitigated Negative Declaration for the proposed project in Merced County. Caltrans is the lead agency under the California Environmental Quality Act (CEQA). The document tells you why the project is being proposed, what alternatives have been considered for the project, how the existing environment could be affected by the project, the potential impacts of each of the alternatives, and the proposed avoidance, minimization, and/or mitigation measures. The Initial Study with Proposed Mitigated Negative Declaration circulated to the public for 30 days between May 31, 2019, and June 29, 2019. Comments received during this period are included in Appendix C. If you would like a printed version or CD of this document, contact Cassidy Ellis by phone at 559-445-6481 or by email at [email protected]. The document can also be downloaded at the following website: https://dot.ca.gov/d10/projects.html Printing this document: To save paper, this document has been set up for two- sided printing (to print the front and back of a page). Blank pages occur where needed throughout the document to maintain proper layout of the chapters and appendices. For individuals with sensory disabilities, this document can be made available in Braille, in large print, on audiocassette, or on computer disk. -

Cramvernal Pool Endemics-Final.Pdf
Vernal Pool Systems and Individual Vernal Pools Version 6.1 APPENDIX 1 Vernal Pool Endemic Plant List Use this list to determine if a species is a vernal pool endemic Bsed on Appendix C from: T. Keeler-Wolf, D.R. Elam, K. Lewis, S.A. Flint. 1998. California Vernal Pool Assessment Preliminary Report. State of California, The Resources Agency, Department of Fish and Game. 161 pp. www.dfg.ca.gov/biogeodata/wetlands/pdfs/VernalPoolAssessmentPreliminaryReport.pdf May 2013 ! CRAM%Vernal%Pool%Endemic%Plants%List May%2013 Scientific%Name Family Genus Species infraspecific_rank %infraspecific_epithet Agrostis(elliottiana POACEAE Agrostis elliottiana Agrostis(hendersonii POACEAE Agrostis hendersonii Agrostis(microphylla POACEAE Agrostis microphylla Alopecurus(carolinianus POACEAE Alopecurus carolinianus Alopecurus(saccatus POACEAE Alopecurus saccatus Anagallis(minima MYRSINACEAE Anagallis minima Astragalus(tener(var.(ferrisiae FABACEAE Astragalus tener var. ferrisiae Astragalus(tener(var.(tener FABACEAE Astragalus tener var. tener Atriplex(cordulata CHENOPODIACEAE Atriplex cordulata Atriplex(cordulata(var.(cordulata CHENOPODIACEAE Atriplex cordulata var. cordulata Atriplex(cordulata(var.(erecticaulis CHENOPODIACEAE Atriplex cordulata var. erecticaulis Atriplex(depressa CHENOPODIACEAE Atriplex depressa Atriplex(minuscula CHENOPODIACEAE Atriplex minuscula Atriplex(parishii CHENOPODIACEAE Atriplex parishii Atriplex(persistens CHENOPODIACEAE Atriplex persistens Atriplex(subtilis CHENOPODIACEAE Atriplex subtilis Blennosperma(bakeri ASTERACEAE Blennosperma -

California Wetlands
VOL. 46, NO.2 FREMONTIA JOURNAL OF THE CALIFORNIA NATIVE PLANT SOCIETY California Wetlands 1 California Native Plant Society CNPS, 2707 K Street, Suite 1; Sacramento, CA 95816-5130 Phone: (916) 447-2677 • Fax: (916) 447-2727 FREMONTIA www.cnps.org • [email protected] VOL. 46, NO. 2, November 2018 Memberships Copyright © 2018 Members receive many benefits, including a subscription toFremontia California Native Plant Society and the CNPS Bulletin. Look for more on inside back cover. ISSN 0092-1793 (print) Mariposa Lily.............................$1,500 Family..............................................$75 ISSN 2572-6870 (online) Benefactor....................................$600 International or library...................$75 Patron............................................$300 Individual................................$45 Gordon Leppig, Editor Plant lover.....................................$100 Student/retired..........................$25 Michael Kauffmann, Editor & Designer Corporate/Organizational 10+ Employees.........................$2,500 4-6 Employees..............................$500 7-10 Employees.........................$1,000 1-3 Employees............................$150 Staff & Contractors Dan Gluesenkamp: Executive Director Elizabeth Kubey: Outreach Coordinator Our mission is to conserve California’s Alfredo Arredondo: Legislative Analyst Sydney Magner: Asst. Vegetation Ecologist native plants and their natural habitats, Christopher Brown: Membership & Sales David Magney: Rare Plant Program Manager and increase understanding, -
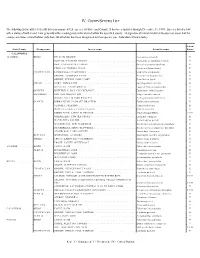
Iv. County/Species List
IV. COUNTY/SPECIES LIST The following list identifies federally listed or proposed U.S. species by State and County. It has been updated through December 31, 1999. Species listed below with a status of both E and T are generally either endangered or threatened within the specified county. Designation of critical habitat (CH) does not mean that the county constitutes critical habitat, only that critical habitat has been designated for that species (see Addendum A Instructions). Action/ State/County Group name Inverse name Scientific name Status CALIFORNIA ALAMEDA ......... BIRDS ....... PELICAN, BROWN ........................................ Pelicanus occidentalis E PLOVER, WESTERN SNOWY ................................ Charadrius alexandrinus nivosus T RAIL, CALIFORNIA CLAPPER ............................... Rallus longirostris obsoletus E TERN, CALIFORNIA LEAST ................................. Sterna antillarum browni E CRUSTACEAN . LINDERIELLA, CALIFORNIA ................................ Linderiella occidentalis E SHRIMP, LONGHORN FAIRY ................................ Branchinecta longiantenna E SHRIMP, VERNAL POOL FAIRY .............................. Branchinecta lynchi T FISHES ...... GOBY, TIDEWATER ...................................... Eucyclogobius newberryi E SPLITTAIL, SACRAMENTO ................................. Pogonichthys macrolepidotus T INSECTS ..... BUTTERFLY, BAY CHECKERSPOT ........................... Euphydryas editha bayensis T MAMMALS ... FOX, SAN JOAQUIN KIT .................................... Vulpes macrotis -

A Checklist of Vascular Plants Endemic to California
Humboldt State University Digital Commons @ Humboldt State University Botanical Studies Open Educational Resources and Data 3-2020 A Checklist of Vascular Plants Endemic to California James P. Smith Jr Humboldt State University, [email protected] Follow this and additional works at: https://digitalcommons.humboldt.edu/botany_jps Part of the Botany Commons Recommended Citation Smith, James P. Jr, "A Checklist of Vascular Plants Endemic to California" (2020). Botanical Studies. 42. https://digitalcommons.humboldt.edu/botany_jps/42 This Flora of California is brought to you for free and open access by the Open Educational Resources and Data at Digital Commons @ Humboldt State University. It has been accepted for inclusion in Botanical Studies by an authorized administrator of Digital Commons @ Humboldt State University. For more information, please contact [email protected]. A LIST OF THE VASCULAR PLANTS ENDEMIC TO CALIFORNIA Compiled By James P. Smith, Jr. Professor Emeritus of Botany Department of Biological Sciences Humboldt State University Arcata, California 13 February 2020 CONTENTS Willis Jepson (1923-1925) recognized that the assemblage of plants that characterized our flora excludes the desert province of southwest California Introduction. 1 and extends beyond its political boundaries to include An Overview. 2 southwestern Oregon, a small portion of western Endemic Genera . 2 Nevada, and the northern portion of Baja California, Almost Endemic Genera . 3 Mexico. This expanded region became known as the California Floristic Province (CFP). Keep in mind that List of Endemic Plants . 4 not all plants endemic to California lie within the CFP Plants Endemic to a Single County or Island 24 and others that are endemic to the CFP are not County and Channel Island Abbreviations .