Sturgeon and Paddlefish Life History and Management: Experts' Knowledge and Beliefs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
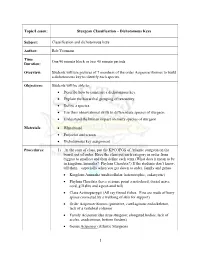
Sturgeon Classification – Dichotomous Keys Subject
Topic/Lesson: Sturgeon Classification – Dichotomous Keys Subject: Classification and dichotomous keys Author: Rob Yeomans Time One 90 minute block or two 45 minute periods Duration: Overview: Students will use pictures of 7 members of the order Acipenseriformes to build a dichotomous key to identify each species. Objectives: Students will be able to: • Describe how to construct a dichotomous key. • Explain the hierarchal grouping of taxonomy. • Define a species. • Use their observational skills to differentiate species of sturgeon. • Understand the human impact on many species of sturgeon. Materials: • Whiteboard • Projector and screen • Dichotomous key assignment Procedures: 1) At the start of class, put the KPCOFGS of Atlantic sturgeon on the board, out of order. Have the class put each category in order from biggest to smallest and then define each term (What does it mean to be in kingdom Animalia? Phylum Chordata?) If the students don’t know, tell them—especially when you get down to order, family and genus. • Kingdom Animalia (multicellular, heterotrophic, eukaryotic) • Phylum Chordata (have at some point a notochord, dorsal nerve cord, gill slits and a post-anal tail) • Class Actinopterygii (All ray finned fishes. Fins are made of bony spines connected by a webbing of skin for support) • Order Acipenseriformes (primitive, cartilaginous endoskeleton, lack of a vertebral column) • Family Acipenseridae (true sturgeon; elongated bodies, lack of scales, anadromous, bottom feeders) • Genus Acipenser (Atlantic Sturgeon) 1 • Species oxyrinchus Comment to the class that there are actually two subspecies of oxyrinchus. Acipenser oxyrinchus oxyrinchus is the Atlantic sturgeon and Acipenser oxyrinchus desotoi is the Gulf sturgeon 2) Reinforce the fact that sturgeon are a primitive fish and fossils have been found dating back 144-65 million years ago. -

Yangtze Sturgeon (Acipenser Dabryanus) - Sturgeons
Pond Life - Yangtze Sturgeon (Acipenser dabryanus) - Sturgeons http://www.pond-life.me.uk/sturgeon/acipenserdabryanus.php Search Pond Life... Home Sturgeons Koi Other Fish Fish Health Ponds Plants Forums Contents Yangtze Sturgeon (Acipenser dabryanus) Home Sturgeons Acipenseriformes Sturgeon Food & Feeding Sturgeon Care Sheet Sturgeon Guide Sturgeon Species List Adriatic Sturgeon Alabama Sturgeon Amu Darya Sturgeon Amur Sturgeon Atlantic Sturgeon Beluga Sturgeon Chinese Paddlefish Chinese Sturgeon Yangtze Sturgeon (Acipenser dabryanus) photo from the website of CAFS Common Sturgeon (http://zzzy.fishinfo.cn/) Diamond Sturgeon Dwarf Sturgeon by Karen Paul Green Sturgeon Description: The Yangtze Sturgeon (Acipenser dabryanus) has 8-13 dorsal scutes, 26-39 lateral Gulf Sturgeon scutes, 9-13 ventral scutes, 44-57 dorsal fin rays and 25-36 anal fin rays. Colouration ranges from Kaluga Sturgeon dark grey to brown-grey on the back to white on the ventral side. The body is rough because it is Lake Sturgeon covered with small pointed denticles. The four barbels are located closer to the mouth than the end Paddlefish of the snout. The Yangtze Sturgeon can reach 1.3 meters in length and a weight of 16kg. Pallid Sturgeon Persian Sturgeon Sakhalin Sturgeon Ship Sturgeon Shortnose Sturgeon Shovelnose Sturgeon Siberian Sturgeon Stellate Sturgeon Sterlet Syr Darya Sturgeon White Sturgeon Yangtze Sturgeon Sturgeon Videos Koi Other Fish Fish Health Yangtze Sturgeon (Acipenser dabryanus) photo from the website of CAFS Ponds (http://zzzy.fishinfo.cn/) Plants Forums Wild Distribution: Asia; restricted to the upper and middle reaches of the Yangtze River system, Search rarely seen below the Gezhouba Dam. The Yangtze Sturgeon is a potamodromous (freshwater only) species. -

Is There Evidence That the Chinese Paddlefish (Psephurus Gladius) Still
Journal of Applied Ichthyology J. Appl. Ichthyol. 25 (Suppl. 2) (2009), 95–99 Received: September 19, 2008 Ó 2009 The Authors Accepted: December 22, 2008 Journal compilation Ó 2009 Blackwell Verlag GmbH doi: 10.1111/j.1439-0426.2009.01268.x ISSN 0175–8659 Is there evidence that the Chinese paddlefish (Psephurus gladius) still survives in the upper Yangtze River? Concerns inferred from hydroacoustic and capture surveys, 2006–2008 By H. Zhang1,2,Q.W.Wei1,2,3,H.Du2,3, L. Shen2,3,Y.H.Li1,2 and Y. Zhao1,2 1College of Fishery, Huazhong Agricultural University, Wuhan, Hubei Province, China; 2Key Laboratory of Freshwater Biodiversity Conservation and Utilization, Ministry of Agriculture of China, Yangtze River Fisheries Research Institute, Chinese Academy of Fisheries Science, Jingzhou, Hubei Province, China; 3Freshwater Fisheries Research Center, Chinese Academy of Fisheries Science, Wuxi, Jiangsu Province, China Summary to save rare and endemic fish species (Fu et al., 2003). Due to the rarity of Chinese paddlefish (Psephurus gladius), very P. gladius has been considered as one of the most important little information is available on this species. In order to save ÔflagshipÕ species in conservation, with an integrated rescue this critically endangered giant species, four large-range hydro- programme initiated in 2005. This extensive programme acoustic surveys (using an echo sounder with a 199 kHz, 6.8° includes studies on forage fishes and habitat investigations, split-beam transducer) and capture surveys (by drift nets and captive breeding programmes and propagation for release, setlines) were conducted in the upper Yangtze River during preservation of genetic resources, and even cloning. -

Caviar and Conservation
Caviar and Conservation Status, Management, and Trade of North American Sturgeon and Paddlefish Douglas F.Williamson May 2003 TRAFFIC North America World Wildlife Fund 1250 24th Street NW Washington DC 20037 Visit www.traffic.org for an electronic edition of this report, and for more information about TRAFFIC North America. © 2003 WWF. All rights reserved by World Wildlife Fund, Inc. All material appearing in this publication is copyrighted and may be reproduced with permission. Any reproduction, in full or in part, of this publication must credit TRAFFIC North America. The views of the author expressed in this publication do not necessarily reflect those of the TRAFFIC Network, World Wildlife Fund (WWF), or IUCN-The World Conservation Union. The designation of geographical entities in this publication and the presentation of the material do not imply the expression of any opinion whatsoever on the part of TRAFFIC or its supporting organizations concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The TRAFFIC symbol copyright and Registered Trademark ownership are held by WWF. TRAFFIC is a joint program of WWF and IUCN. Suggested citation: Williamson, D. F. 2003. Caviar and Conservation: Status, Management and Trade of North American Sturgeon and Paddlefish. TRAFFIC North America. Washington D.C.: World Wildlife Fund. Front cover photograph of a lake sturgeon (Acipenser fulvescens) by Richard T. Bryant, courtesy of the Tennessee Aquarium. Back cover photograph of a paddlefish (Polyodon spathula) by Richard T. Bryant, courtesy of the Tennessee Aquarium. TABLE OF CONTENTS Preface . -

Ecological Effects of the First Dam on Yangtze Main Stream and Future Conservation Recommendations: a Review of the Past 60 Years
Zhang et al.: Ecological effects of the first dam on Yangtze main stream - 2081 - ECOLOGICAL EFFECTS OF THE FIRST DAM ON YANGTZE MAIN STREAM AND FUTURE CONSERVATION RECOMMENDATIONS: A REVIEW OF THE PAST 60 YEARS ZHANG, H.1 – LI, J. Y.1 – WU, J. M.1 – WANG, C. Y.1 – DU, H.1 – WEI, Q. W.1* – KANG, M.2* 1Key Laboratory of Freshwater Biodiversity Conservation, Ministry of Agriculture of China; Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan, Hubei Province, P. R. China (phone: +86-27-8178-0118; fax: +86-27-8178-0118) 2Department of Maritime Police and Production System / The Institute of Marine Industry, Gyeongsang National University, Cheondaegukchi-Gil 38, Tongyeong-si, Gyeongsangnam-do, 53064, South Korea (phone: +82-55-772-9187; fax: +82-55-772-9189) *Corresponding authors e-mail: [email protected]; [email protected] (Received 21st Jul 2017; accepted 27th Oct 2017) Abstract. The Gezhouba Dam was the first and lowermost dam on the major stem of the Yangtze River. Up to now, the dam has been operating for more than 35 years. The time period was a fast economic development stage in the Yangtze basin. Therefore, the entire Yangtze aquatic ecosystem has been highly affected by various anthropogenic activities. Especially, the fish population and distribution in the Yangtze River have been largely altered. This study reviews the ecological effects of the Gezhouba Dam to the Yangtze aquatic biodiversity for the past 60 years based on literatures. It was concluded that the pre-assessment of the Gezhouba Dam on Yangtze fishes in 1970s was appropriate. -

SAVING STURGEONS a Global Report on Their Status and Suggested Conservation Strategy the Report Is a Joint Effort of the WWF Network
SAVING STURGEONS A global report on their status and suggested conservation strategy The report is a joint effort of the WWF network. It was written by Ralf Reinartz (consultant) and Polina Slavche- va (WWF Danube-Carpathian Programme) and coordinated by Polina Slavcheva. Special thanks to Esther Blom (WWF-Neth- erlands), Judy Takats (WWF-US), Lin Cheng and Jinyu Lei (WWF-China), Alexander Moiseev (WWF-Russia), Vesselina Kavrakova, Stoyan Mihov and Ekaterina Voynova (WWF DCP-Bulgaria), Cristina Munteanu and George Caracas (WWF DCP-Romania) and Samantha Ampel for their contributions. WWF would like to acknowledge TRAFFIC and the IUCN for their support of WWF’s sturgeon conservation work. Design and production: Marieta Vasileva, Taralej Ltd. © Front cover photo: Zdravko Yonchev WWF is one of the world’s largest and most experienced independent conservation organisations, with over 5 million supporters and a global network active in more than 100 coun- tries. WWF’s mission is to stop the degradation of the planet’s natural environment and to build a future in which humans live in harmony with nature, by conserving the world’s biological diversity, ensuring that the use of renewable natural resources is sustainable, and promoting the reduction of pollution and wasteful consumption. TABLE OF CONTENTS Foreword 5 PART I. THE EXTRAORDINARY STURGEONS 6 Some extraordinary facts and fi gures on sturgeons 8 Sturgeons in culture and history 11 Global status today 13 Commercial importance 14 PART II. WHY STURGEONS ARE THREATENED 16 Overexploitation 18 Loss of migration routes and suitable habitat 19 Genetic factors 19 PART III. POPULATIONS ON THE BRINK – A JOURNEY TO STURGEON REGIONS AND RIVERS 22 The Northeastern Pacifi c 25 The Great Lakes, Hudson Bay & St. -

Sturgeon and Paddlefish Life History and Management: Experts’ Knowledge and Beliefs
Received: 18 May 2017 | Accepted: 16 June 2017 DOI: 10.1111/jai.13563 STURGEON PAPER Sturgeon and paddlefish life history and management: Experts’ knowledge and beliefs I. Jarić1,2,3 | C. Riepe1,4 | J. Gessner1 1Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Berlin, Germany Summary 2Biology Centre of the Czech Academy of Although sturgeons and paddlefish represent a highly endangered species group, Sciences, Institute of Hydrobiology, České knowledge regarding their ecology, life history and management and restoration Budějovice, Czech Republic measures still reveals substantial gaps and uncertainties. As such, it was hypothesized 3Institute for Multidisciplinary Research, University of Belgrade, Belgrade, that the information on overall experiences and perspectives of researchers working Serbia in the field would provide a good indication of the state of knowledge, and help iden- 4Thünen-Institute of Baltic Sea Fisheries, Rostock, Germany tify areas for further investigation. To gain in- depth insight into their perspectives on a variety of topics related to sturgeon life history, management and conservation, as Correspondence Ivan Jarić, Biology Centre of the Czech well as opinions on issues characterized by a lack of data, a worldwide online survey Academy of Sciences, Institute of was conducted among scientists specializing in sturgeon and paddlefish research. Hydrobiology, České Budějovice, Czech Republic. With a total of 277 respondents, the response rate was good (40.3% of those invited). Email: [email protected]; [email protected] The survey results indicated a high level of uncertainty with regard to various aspects of sturgeon life history, such as spawning migrations, reproduction and senescence. Responses differed largely with the origin and the research focus of the participants. -

Abstract 1 the Paddlefish Polyodon Spathula Is a Ram Suspension
1 Abstract 2 The Paddlefish Polyodon spathula is a ram suspension feeding planktivore that resides in fresh 3 water habitats. While there are numerous hypotheses about the external morphology of the Paddlefish, 4 none truly support a definite finding. 5 Context and Content 6 The Paddlefish Polyodon spathula is classified as the following: Kingdom: Animalia, Phylum: 7 Chordata, Class: Actinopterygii, Order: Acipenseriformes, Family: Polyodontidae, Genus: Polyodon, 8 Species: spathula (Ross and Brenneman 2001, Starnes 2016). The order Acipenseriformes includes the 9 sturgeons (family Acipenseridae) and the paddlefishes (family Polyodontidae; Ross and Brenneman 10 2001). The Chinese Paddlefish, Psephurus gladius, is the only other species in the family Polyodontidae 11 (Wilkens and Hofmann 2007) and resides exclusively in China’s Yangtze River (Ross and Brenneman 12 2001). There are a variety of common names associated with the species such as Paddlefish, Spadefish, 13 Spoonbill Catfish, or Duck-Billed Catfish (Ross and Brenneman 2001). Walbaum portrayed the Paddlefish 14 as a shark in 1792; but was corrected by Lacepedѐ in 1797, who classified Paddlefish in the genus 15 Polyodon (McKinley 1984). 16 General Characteristics 17 Paddlefish are an ancient species and exhibit less derived traits. They are typically characterized 18 by the presence of heterocercal caudal fins and the absence of teeth in adults (Ross and Brenneman 19 2001). Polyodon means ‘many teeth’, while Adult Paddlefish do not have teeth, but do have many gill 20 rakers which they use to filter zooplankton from the water column (Ross and Brenneman 2001) and 21 spathula means ‘flat piece’, referring to the long snout, or rostrum, which makes up around one third of 22 the total body length of adults (Wilkens and Hofmann 2007). -

Universal Caviar Labelling Requirements
UNIVERSAL CAVIAR LABELLING REQUIREMENTS All sturgeon caviar BACKGROUND containers in domestic and international trade have to All species of sturgeon and paddlefish have been listed in the Appendices of the bear a non-reusable label Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) since 1998. Two species—the Common or Baltic Sturgeon Acipenser sturio containing details about the and the Shortnose Sturgeon Acipenser brevirostrum—are listed in Appendix I of the source and the country of Convention, which prohibits any international commercial trade. All other species origin of the caviar. are listed in Appendix II, whereby international trade is regulated by governments Governments around the through a system of permits. world have agreed to a universal caviar labelling With the aim of facilitating the legal caviar trade around the world and allowing the system with the aim of easy identification of the source and origin of caviar, governments agreed at a CITES ensuring that all caviar meeting in 2000 to introduce a standardized labelling system for all caviar exports. Two years later, they extended the labelling requirements and agreed that all caviar entering the market is from containers in trade, whether imported, exported, re-exported or in domestic legal sources. The caviar markets, should bear a label that would contain a specific set of information, including labelling system helps the country of origin and the year of harvest, to allow identification of the source of governments, traders and the caviar. Governments agreed that, as of January 2004, they would only accept consumers in distinguishing imported caviar shipments marked according to the labelling guidelines. -

The Basics of Legal Caviar Trade
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: WWF Studien, Broschüren und sonstige Druckmedien Jahr/Year: 2014 Band/Volume: 48_2014 Autor(en)/Author(s): Anonymus Artikel/Article: Sturgeons and caviar The basics of legal caviar trade. 1-2 ©Umweltverband WWF Österreich; download unter www.zobodat.at CITES SPECIES CODES FOR ALL STURGEON SPECIES CITES SPECIES CODE SCIENTIFIC NAME ENGLISH NAME BAE Acipenser baerii Siberian sturgeon BAI Acipenser baerii baicalensis Baikal sturgeon BVI Acipenser brevirostrum Shortnose Sturgeon DAB Acipenser dabryanus Yangtze Sturgeon FUL Acipenser fulvescens Lake Sturgeon GUE Acipenser gueldenstaedtii Russian Sturgeon MED Acipenser medirostris Green Sturgeon MIK Acipenser mikadoi Sakhalin Sturgeon NAC Acipenser naccarii Adriatic Sturgeon NUD Acipenser nudiventris Ship Sturgeon OXY Acipenser oxyrhynchus Atlantic Sturgeon DES Acipenser oxyrhynchus desotoi Gulf Sturgeon PER Acipenser persicus Persian Sturgeon RUT Acipenser ruthenus Sterlet SCH Acipenser schrenckii Amur Sturgeon SIN Acipenser sinensis Chinese Sturgeon STE Acipenser stellatus Stellate Sturgeon The basics of legal STU Acipenser sturio European Sturgeon TRA Acipenser transmontanus White Sturgeon DAU Huso dauricus Kaluga Sturgeon caviar trade HUS Huso huso Beluga Sturgeon SPA Polyodon spathula American Paddlefish © WWF Austria / Jutta Jahrl © WWF GLA Psephurus gladius Chinese Paddlefish FED Pseudoscaphirhynchus fedtschenkoi Syr-darya Shovelnose Sturgeon HER Pseudoscaphirhynchus hermanni Small Amu-dar Shovelnose or Dwarf Sturgeon KAU Pseudoscaphirhynchus kaufmanni Large Amu-dar or False Shovelnose Sturgeon ALB Scaphirhynchus albus Pallid Sturgeon PLA Scaphirhynchus platorynchus Shovelnose Sturgeon SUS Scaphirhynchus suttkusi Alabama Sturgeon MIX Mixed species (for ‘pressed’ caviar exclusively) YYYxXXX Hybrid specimens: code for the species of the male x code for the species of the female ©Umweltverband WWF Österreich; download unter www.zobodat.at. -

Chinese Paddlefish, Chinese Swordfish, White Sturgeon Finnish: Miekkasampi French: German: Polish: Wioslonos Chinski Spanish
263 Proposal II / 31 PROPOSAL FOR INCLUSION OF SPECIES ON THE APPENDICES OF THE CONVENTION ON THE CONSERVATION OF MIGRATORY SPECIES OF WILD ANIMALS PROPOSAL: Inclusion of the following species of Psephurus gladius in Appendix II of the Convention on the Conservation of Migratory Species of Wild Animals (CMS): B. PROPONENT: Federal Republic of Germany C. SUPPORTING STATEMENT 1. Taxon 1.1 Classis: Actinopterygii 1.2 Ordo: Acipenseriformes 1.3 Familia: Polyodontidae 1.4 Species: Psephurus gladius (Martens, 1862) 1.5 Common names Chinese: Yin yui, Wei, Chin yui English: Chinese paddlefish, Chinese swordfish, White sturgeon Finnish: Miekkasampi French: German: Polish: Wioslonos chinski Spanish: 2. Biological data 2.1 Distribution Psephurus gladius is endemic to the Yangtze River (Chang Jiang) and its attached tributaries, including the rivers of Tuojiang, Mingijang, Jialimjiang, Qiuantangjiang and Yongjiang, and the lakes of Dongting and Poayang (Liu and Zeng, 1988; Liu et al., 1995). In the past, the species was most commonly found in the trunk of the Yangtze River and only rarely encountered in the tributaries (Wei et al, 1997). Mature adults of Psephurus gladius are reported to migrate to sea and are historically documented in the East China Sea, The Yellow Sea and The Yellow River (Liu and Zeng, 1988). Furthermore, individuals are sometimes accidentally brought in form the sea by high spring tides, so that they are rarely discovered in the lower reaches of the Qiantang and Yangjiang rivers of Zhejiang Province (Liu and Zeng, 1988). 2.2 Population According to Liu (1995) the entire population of Psephurus gladius is decreasing, mainly since 1976 due to overfishing and pollution. -

Sturgeon Hatchery Manual
FAO ISSN 2070-7010 FISHERIES AND 558 AQUACULTURE TECHNICAL PAPER 558 STURGEON HATCHERY MANUAL This Sturgeon Hatchery Manual includes the latest available scientific research findings and experiences and compiles advice given in earlier manuals and handbooks on sturgeon culture and reproduction practices. This document was prepared in response HATC STURGEON to numerous requests for practical guidance on this subject from the Central Asian and Caucasus region to the Food and Agriculture Organization of the United Nations (FAO). This manual is targeted particularly at sturgeon farmers, sturgeon hatchery operators, hatchery technicians, and fisheries and aquaculture managers involved in sturgeon aquaculture development and the restocking and rehabilitation of sturgeon populations in the countries around the basins of the Black and Caspian seas. It aims to provide a practical handbook of modern sturgeon hatchery practices and management. The H manual is available in the English, Russian and Turkish languages. MANUAL ERY FAO FAO Sturgeon Hatchery FISHERIES AND Manual AQUACULTURE TECHNICAL PAPER 558 Prepared by Mikhail S. Chebanov FAO Consultant Krasnodar, Russia and Elena V. Galich Krasnodar, Russia FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Ankara, 2013 Reprinted 2013 The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned.