Sturgeon and Paddlefish Life History and Management: Experts’ Knowledge and Beliefs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Federal Register/Vol. 70, No. 189/Friday, September 30, 2005
57316 Federal Register / Vol. 70, No. 189 / Friday, September 30, 2005 / Notices Dated: September 22, 2005. 2. Chairman’s Report. ADDRESSES: Caspian Sea littoral states Mark R. Johnston, 3. Executive Director’s Report. wishing to provide information that may Director, Office of Special Needs Assistance 4. Financial Budget. allow us to lift this trade suspension Programs. 5. Public Input. may submit it by any one of several [FR Doc. 05–19298 Filed 9–29–05; 8:45 am] It is anticipated that about twenty-five methods: people will be able to attend the session BILLING CODE 4210–29–M 1. You may submit written in addition to the Commission information to Robert R. Gabel, Chief, members. Division of Scientific Authority, U.S. Interested persons may make oral or INTER-AMERICAN FOUNDATION Fish and Wildlife Service, 4401 North written presentations to the Commission Fairfax Drive, Room 750, Arlington, or file written statements. Such requests Sunshine Act Meeting; Agenda for Virginia 22203. should be made prior to the meeting to: board of Directors’ Meeting 2. You may hand-deliver written Larry Gall, Interim Executive Director, information to the Division of Scientific October 14, 2005, 9:30 a.m.–3 p.m. John H. Chafee, Blackstone River Valley Authority, at the above address, or fax The meeting will be held at the Inter- National Heritage Corridor Commission, your comments to (703) 358–2276. American Foundation, 901 N. Stuart One Depot Square, Woonsocket, RI 3. You may send information by Street, 10th Floor, Arlington, Virginia 02895, Tel.: (401) 762–0250. electronic mail (e-mail) to 22203. -

First Verified Occurrence of the Shortnose Sturgeon (Acipenser Brevirostrum) in the James River, Virginia
19 6 Abstract—The shortnose sturgeon (Acipenser brevirostrum) is an en- First verified occurrence of the shortnose dangered species of fish that inhab- sturgeon (Acipenser brevirostrum) its the continental slope of the At- lantic Ocean from New Brunswick, in the James River, Virginia Canada, to Florida. This species has not been documented previously in the freshwater portion of any river Matthew Balazik of the Chesapeake Bay, except in the Potomac River. On 13 March 2016, a Email address for author: [email protected] shortnose sturgeon was captured in the freshwater portion of the James Center for Environmental Studies River at river kilometer 48. The Virginia Commonwealth University fish had a fork length of about 75 1000 West Cary Street cm and was likely mature. Genetic Richmond, Virginia 23284 analysis confirmed the fish was a shortnose sturgeon and was assigned to the Chesapeake Bay–Delaware population segment. Regardless of whether this shortnose sturgeon was part of a remnant Chesapeake The shortnose sturgeon (Acipenser ture of shortnose sturgeon, as well Bay population or whether its cap- brevirostrum) is an amphidromous (Spells1; Welsh et al., 2002; Mangold ture there is an indicator of an ex- sturgeon reported to inhabit the con- et al.2). Only one genetically verified pansion of range from the Delaware tinental slope of the Atlantic Ocean shortnose sturgeon was collected in River by way of the Chesapeake and from New Brunswick, Canada, to Virginia waters as part of these pro- Delaware Canal, dedicated research Florida (Gruchy and Parker, 1980; grams (Spells1; Welsh et al., 2002). is needed to determine the status of Dadswell et al., 1984; Kynard, 1997). -

Atlantic Sturgeon
Species Profile: Atlantic Sturgeon Benchmark Assessment Indicates Signs of a Slow Recovery Species Snapshot Though Challenges Towards Sustainability Remain Introduction Atlantic Sturgeon For almost 30 years, the 15 Atlantic coastal states have worked together to effectively Acipenser oxyrhynchus oxyrhynchus manage Atlantic sturgeon throughout its range from Maine to Florida. Recognizing both the importance of this ancient species and the dire status of the population, the states Management Unit: Maine to Florida implemented a 40-year coastwide moratorium on harvest through Amendment I to the Interesting Facts Atlantic Sturgeon FMP in 1998 with the goal of restoring the population of this once thriving • The species name 'oxyrhynchus' means sharp fishery. Since then, the states have invested considerable resources to research the species’ snout. biology and life history. Despite the strong conservation efforts taken, Atlantic sturgeon was added to the Endangered Species List in February 2012. A coastwide benchmark stock • Sturgeon were a key food source for U.S. settlers assessment completed by the Commission in the fall of 2017 concluded that the population along the Atlantic coast. remains depleted at the coastwide and distinct population segment (DPS) levels relative • In the late 1800s, fishing for sturgeon eggs (to to historic abundance, although the population appears to be recovering slowly since sell as caviar) attracted so many people, the trend implementation of the 1998 moratorium. was referred to as the “Black Gold Rush.” • Atlantic sturgeon are river-specific, returning to Life History their natal rivers to spawn. Atlantic sturgeon (Acipenser oxyrhynchus oxyrhynchus) are ancient fish, dating back to at least • Rather than having true scales, Atlantic sturgeon the late Cretaceous Period (66-100 million years ago). -

Pan-European Action Plan for Sturgeons
Strasbourg, 30 November 2018 T-PVS/Inf(2018)6 [Inf06e_2018.docx] CONVENTION ON THE CONSERVATION OF EUROPEAN WILDLIFE AND NATURAL HABITATS Standing Committee 38th meeting Strasbourg, 27-30 November 2018 PAN-EUROPEAN ACTION PLAN FOR STURGEONS Document prepared by the World Sturgeon Conservation Society and WWF This document will not be distributed at the meeting. Please bring this copy. Ce document ne sera plus distribué en réunion. Prière de vous munir de cet exemplaire. T-PVS/Inf(2018)6 - 2 - Pan-European Action Plan for Sturgeons Multi Species Action Plan for the: Russian sturgeon complex (Acipenser gueldenstaedtii, A. persicus-colchicus), Adriatic sturgeon (Acipenser naccarii), Ship sturgeon (Acipenser nudiventris), Atlantic/Baltic sturgeon, (Acipenser oxyrinchus), Sterlet (Acipenser ruthenus), Stellate sturgeon (Acipenser stellatus), European/Common sturgeon (Acipenser sturio), and Beluga (Huso huso). Geographical Scope: European Union and neighbouring countries with shared basins such as the Black Sea, Mediterranean, North Eastern Atlantic Ocean, North Sea and Baltic Sea Intended Lifespan of Plan: 2019 – 2029 Russian Sturgeon Adriatic Sturgeon Ship Sturgeon Atlantic or Baltic complex Sturgeon Sterlet Stellate Sturgeon Beluga European/Common Sturgeon © M. Roggo f. A. sturio; © Thomas Friedrich for all others Supported by - 3 - T-PVS/Inf(2018)6 GEOGRAPHICAL SCOPE: The Action Plan in general addresses the entire Bern Convention scope (51 Contracting Parties, including the European Union) and in particular the countries with shared sturgeon waters in Europe. As such, it focuses primarily on the sea basins in Europe: Black Sea, Mediterranean, North-East Atlantic, North Sea, Baltic Sea, and the main rivers with relevant current or historic sturgeon populations (see Table 2). -

Life History and Population Dynamics of Lake Sturgeon
LIFE HISTORY AND POPULATION DYNAMICS OF LAKE STURGEON, Acipenser fulvescens, IN THE MUSKEGON RIVER, MICHIGAN by Paul Joseph Vecsei (Under the direction of Douglas L. Peterson) ABSTRACT The lake sturgeon was once abundant throughout Lake Michigan with an estimated 11 million fish prior to human exploitation. By the early 1900s, however, most populations had been decimated by severe over-fishing and habitat degradation. Despite recent interests in restoring the species in Lake Michigan, little is known about the current status of remnant populations. The primary objectives of this study were to estimate annual spawning stock abundance and to identify potential spawning habitat for lake sturgeon on the Muskegon River, Michigan. To capture adult lake sturgeon, I used large-mesh, bottom-set gill nets deployed at the mouth of the Muskegon River from mid- March through May, 2002-2005. Radio telemetry was used to monitor seasonal movements and to identify likely spawning habitats. Sampling for larval lake sturgeon was conducted in May of each year using D-frame drift nets anchored in the mainstream of the river channel. During the 4 years of the study, I expended more than 5000 gill-net hours and captured 59 individual adult lake sturgeon. Larval lake sturgeon were captured in 2 years, suggesting that at least some natural reproduction still occurs. Habitat analysis revealed that the lower Muskegon River likely contains extensive reaches of potential spawning habitat for lake sturgeon. INDEX WORDS: Biology, population dynamics, habitat, lake -

Distribution and Movement of Chinese Sturgeon, Acipenser Sinensis, on the Spawning Ground Located Below the Gezhouba Dam During Spawning Seasons
J. Appl. Ichthyol. 22 (Suppl. 1) (2006), 145-151 Received: January 10, 2006 © 2006 Blackwell Verlag, Berlin Accepted: August 9, 2006 ISSN 0175-8659 Distribution and movement of Chinese sturgeon, Acipenser sinensis, on the spawning ground located below the Gezhouba Dam during spawning seasons By D. Yang1,3, B. Kynard2, Q. Wei1,3, X. Chen1,3, W. Zheng1 and H. Du1,3 1Key Laboratory of Freshwater Fish Germplasm Resources and Biotechnology, the Ministry of Agriculture of PRC. Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Jinzhou 434000, China; 2Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, China; 3S. C. Anadromous Fish Research Center, US Geological Service, MA 01376 USA Summary estuary to begin their upstream spawning migration as maturing adults. The age of females and males at this stage is commonly Sonic tracking of 40 adult pre-spawning Chinese sturgeons (35 above 14 years and 9 years, respectively. Migrating spawners usu- females, 5 males) was done during 8 spawning seasons (1996– ally arrive at the spawning ground about 18 months after starting 2004). Fish spent most of the time close to the dam (within 7 km). the upstream migration. Their gonads develop from stage III into A yearly mean of 64% of the tracking locations of pre-spawners stage IV during the upstream migration (Anonymous. 1988; were in the small area of 1.08 km distance below the dam (cell I-B Chang, 1999). The range of their spawning ground locations before of the tracking grid system). These movements indicate fish con- the construction of the Gezhouba Dam reached from the Maoshui tinue to attempt to move farther upstream to pass the dam. -

Pallid Sturgeon Recovery Plan (Recovery Plan)
PALLID STURGEON coRE VERYPMN Recovery Plan for the Pallid Sturgeon (Scaphirhynchus a/bus) Prepared by the Pallid Sturgeon Recovery Team Principal Authors Mark P. Dryer, Leader U.S. Fish and Wildlife Service Ecological Services 1500 Capitol Avenue Bismarck, ND 58501 and Alan J. Sandvol U.S. Fish and Wildlife Service Fisheries and Federal Aid 1500 Capitol Avenue Bismarck, ND 58501 for Region 6 U.S. Fish and Wildlife Service Denver, Colorado / Approved: Re~5l i rector Date TABLE OF CONTENTS TITLE PAGE RECOVERY BACKGROUND AND STRATEGY iii PALLID STURGEON RECOVERY TEAM iv DISCLAIMER V ACKNOWLEDGEMENTS vi EXECUTIVE SUMMARY vii Part I INTRODUCTION 1 History 1 General Description 1 Historical Distribution and Abundance 3 Present Distribution and Abundance 5 Habitat Preference 5 Current Velocity 7 Turbidity 8 Water Depth 8 Substrate 8 Temperature 8 Life History 8 Reproductive Biology 8 Food and Feeding Habits 9 Age and Growth 10 Reasons for Decline 10 Habitat Loss 10 Commercial Harvest 13 Pall uti on/Contami nants 14 Hybridization 14 Part II RECOVERY 16 Recovery Objectives and Criteria 16 Recovery—Priority Management Areas 16 Recovery Outline 19 24 Recovery Outline Narrative . LITERATURE CITED 42 Part III IMPLEMENTATION SCHEDULE 46 FIGURES NO. PAGE 1. Comparative Diagrams of the Ventral Surface of the Head of Shovelnose Sturgeon and Pallid Sturgeon, Showing Several Measurement Ratios of Value for Identification 2 2. Historic Range of Pallid Sturgeon 4 3. Recent Occurrence of Pallid Sturgeon 6 4. Recovery—Priority Management Areas 18 RECOVERY BACKGROUND AND STRATEGY The pallid sturgeon (Scaphirhynchus albus Forbes and Richardson) was listed as an endangered species on September 6, 1990 (55 FR 36641) pursuant to the Endangered Species Act (Act) of 1973 (16 U.S.C. -

Status of Lake Sturgeon (Acipenser Fulvescens Rafinesque 1817)
Journal of Applied Ichthyology J. Appl. Ichthyol. 32 (Suppl. 1) (2016), 162–190 Received: August 12, 2016 © 2016 Blackwell Verlag GmbH Accepted: October 19, 2016 ISSN 0175–8659 doi: 10.1111/jai.13240 Status of Lake Sturgeon (Acipenser fulvescens Rafinesque 1817) in North America By R. M. Bruch1, T. J. Haxton2, R. Koenigs1, A. Welsh3 and S. J. Kerr2 1Wisconsin Department of Natural Resources, Oshkosh, WI, USA; 2Ontario Ministry of Natural Resources and Forestry, Peterborough, ON, Canada; 3School of Natural Resources, West Virginia University, Morgantown, WV, USA Summary The species Lake Sturgeon (LS) had been assigned at least Lake Sturgeon is a potamodromous, fluvial-dependent spe- 17 different scientific names during the 19th and 20th cen- cies from the family Acipenseridae, and one of the largest turies due to the variation in color and shape displayed by freshwater fishes within its North American range extending the different life stages and populations (Scott and Cross- to the Great lakes, Mississippi River, and Hudson Bay drai- man, 1973). Eventually LS was recognized as one species nages. Like almost all other sturgeon species, Lake Stur- with the scientific designation Acipenser fulvescens (ful- = geon populations throughout its range suffered mass vescens yellowish or tawny) originally proposed by Con- – declines or extirpation in the late 1800s into the early stantine Samuel Rafinesque (1783 1840). The common name 1900s, due to extensive overexploitation and habitat loss Lake Sturgeon was given due to the abundance of the species and -
Sturgeon Classification – Dichotomous Keys Subject
Topic/Lesson: Sturgeon Classification – Dichotomous Keys Subject: Classification and dichotomous keys Author: Rob Yeomans Time One 90 minute block or two 45 minute periods Duration: Overview: Students will use pictures of 7 members of the order Acipenseriformes to build a dichotomous key to identify each species. Objectives: Students will be able to: • Describe how to construct a dichotomous key. • Explain the hierarchal grouping of taxonomy. • Define a species. • Use their observational skills to differentiate species of sturgeon. • Understand the human impact on many species of sturgeon. Materials: • Whiteboard • Projector and screen • Dichotomous key assignment Procedures: 1) At the start of class, put the KPCOFGS of Atlantic sturgeon on the board, out of order. Have the class put each category in order from biggest to smallest and then define each term (What does it mean to be in kingdom Animalia? Phylum Chordata?) If the students don’t know, tell them—especially when you get down to order, family and genus. • Kingdom Animalia (multicellular, heterotrophic, eukaryotic) • Phylum Chordata (have at some point a notochord, dorsal nerve cord, gill slits and a post-anal tail) • Class Actinopterygii (All ray finned fishes. Fins are made of bony spines connected by a webbing of skin for support) • Order Acipenseriformes (primitive, cartilaginous endoskeleton, lack of a vertebral column) • Family Acipenseridae (true sturgeon; elongated bodies, lack of scales, anadromous, bottom feeders) • Genus Acipenser (Atlantic Sturgeon) 1 • Species oxyrinchus Comment to the class that there are actually two subspecies of oxyrinchus. Acipenser oxyrinchus oxyrinchus is the Atlantic sturgeon and Acipenser oxyrinchus desotoi is the Gulf sturgeon 2) Reinforce the fact that sturgeon are a primitive fish and fossils have been found dating back 144-65 million years ago. -

2012 Wildearth Guardians and Friends of Animals Petition to List
PETITION TO LIST Fifteen Species of Sturgeon UNDER THE U.S. ENDANGERED SPECIES ACT Submitted to the U.S. Secretary of Commerce, Acting through the National Oceanic and Atmospheric Administration and the National Marine Fisheries Service March 8, 2012 Petitioners WildEarth Guardians Friends of Animals 1536 Wynkoop Street, Suite 301 777 Post Road, Suite 205 Denver, Colorado 80202 Darien, Connecticut 06820 303.573.4898 203.656.1522 INTRODUCTION WildEarth Guardians and Friends of Animals hereby petitions the Secretary of Commerce, acting through the National Marine Fisheries Service (NMFS)1 and the National Oceanic and Atmospheric Administration (NOAA) (hereinafter referred as the Secretary), to list fifteen critically endangered sturgeon species as “threatened” or “endangered” under the Endangered Species Act (ESA) (16 U.S.C. § 1531 et seq.). The fifteen petitioned sturgeon species, grouped by geographic region, are: I. Western Europe (1) Acipenser naccarii (Adriatic Sturgeon) (2) Acipenser sturio (Atlantic Sturgeon/Baltic Sturgeon/Common Sturgeon) II. Caspian Sea/Black Sea/Sea of Azov (3) Acipenser gueldenstaedtii (Russian Sturgeon) (4) Acipenser nudiventris (Ship Sturgeon/Bastard Sturgeon/Fringebarbel Sturgeon/Spiny Sturgeon/Thorn Sturgeon) (5) Acipenser persicus (Persian Sturgeon) (6) Acipenser stellatus (Stellate Sturgeon/Star Sturgeon) III. Aral Sea and Tributaries (endemics) (7) Pseudoscaphirhynchus fedtschenkoi (Syr-darya Shovelnose Sturgeon/Syr Darya Sturgeon) (8) Pseudoscaphirhynchus hermanni (Dwarf Sturgeon/Little Amu-Darya Shovelnose/Little Shovelnose Sturgeon/Small Amu-dar Shovelnose Sturgeon) (9) Pseudoscaphirhynchus kaufmanni (False Shovelnose Sturgeon/Amu Darya Shovelnose Sturgeon/Amu Darya Sturgeon/Big Amu Darya Shovelnose/Large Amu-dar Shovelnose Sturgeon/Shovelfish) IV. Amur River Basin/Sea of Japan/Sea of Okhotsk (10) Acipenser mikadoi (Sakhalin Sturgeon) (11) Acipenser schrenckii (Amur Sturgeon) (12) Huso dauricus (Kaluga) V. -
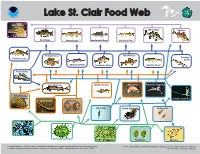
Lake St. Clair Food Web MENT of C
ATMOSPH ND ER A I C C I A N D A M E I C N O I S L T A R N A T O I I O T N A N U E .S C .D R E E PA M RT OM Lake St. Clair Food Web MENT OF C Sea Lamprey White Bass Walleye Rock Bass Muskellunge Smallmouth Bass Northern Pike Pumpkinseed Yellow Perch Rainbow Common Carp Smelt Channel Catfish Freshwater Drum Spottail Shiner Round Goby Lake Sturgeon Mayfly nymphs Gammarus Raptorial waterflea Zebra/Quagga mussels Invasive waterflea Chironomids Mollusks Calanoids Native waterflea Cyclopoids Diatoms Green algae Blue-green algae Flagellates Rotifers Foodweb based on “Impact of exotic invertebrate invaders on food web structure and function in the Great Lakes: NOAA, Great Lakes Environmental Research Laboratory, 4840 S. State Road, Ann Arbor, MI A network analysis approach” by Mason, Krause, and Ulanowicz, 2002 - Modifications for Lake St. Clair, 2009. 734-741-2235 - www.glerl.noaa.gov Lake St. Clair Food Web Sea Lamprey Planktivores/Benthivores Sea lamprey (Petromyzon marinus). An aggressive, non-native parasite that Lake Sturgeon (Acipenser fulvscens). Endangered over most of its historic fastens onto its prey and rasps out a hole with its rough tongue. range. Its diet commonly includes small clams, snails, crayfish, sideswimmers, aquatic insect larvae, algae, and other plant matter. Piscivores (Fish Eaters) Macroinvertebrates White bass (Morone Chrysops). Prefers clear open water in lakes and large rivers. Chironomids/Oligochaetes. Larval insects and worms that live on the lake Visual feeders, uses sight instead of smell to find prey. -

Acipenser Oxyrinchus) Ecological Risk Screening Summary
Atlantic sturgeon (Acipenser oxyrinchus) Ecological Risk Screening Summary U.S. Fish and Wildlife Service, web version – 03/29/2018 Photo: Simon Pierre Barrette. Licensed under CC BY-SA 3.0. Available: https://commons.wikimedia.org/wiki/File:Acipenser_oxyrinchus_PAQ.jpg. 1 Native Range and Status in the United States Native Range From Froese and Pauly (2014): “Western Atlantic: Hamilton River, Labrador, Newfoundland, Canada to northeastern Florida, USA. Occurs occasionally in Bermuda and French Guiana [Robins and Ray 1986]. Northern Gulf of Mexico [Smith 1997]. In Europe: Baltic Sea. Landlocked populations in Lakes Ladoga and Onega (Russia), both now extirpated. Occasionally recorded from Great Britain and North Sea in Elbe drainage [Kottelat and Freyhof 2007]. Recent research revealed that this species existed in the Baltic Sea, but is now extirpated [Ludwig et al. 2002, 2008].” 1 Status in the United States From Froese and Pauly (2014): “USA: native” From NOAA Fisheries (2016): “Historically, Atlantic sturgeon were present in approximately 38 rivers in the United States from St. Croix, ME to the Saint Johns River, FL, of which 35 rivers have been confirmed to have had a historical spawning population. Atlantic sturgeon are currently present in approximately 32 of these rivers, and spawning occurs in at least 20 of them.” GBIF Secretariat (2014) contains a record in California that is assumed to be a misidentification as it is from 1956 and not substantiated elsewhere. Means of Introductions in the United States No records of non-native Acipenser oxyrinchus introductions in the United States were found. Remarks From Froese and Pauly (2014): “Near threatened globally, but extirpated in Europe [Kottelat and Freyhof 2007].