Aves: Accipitriformes) Com Base Em Caracteres Osteológicos
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JNCC Report No. 412 (Addendum): Raptor Distribution Maps
JNCC Report No. 412 (Addendum) Survey of the occurrence and relative abundance of raptors in Guinea subject to international trade Addendum – raptor distribution maps Guy Rondeau, Mohammed Moussa Condeé, Bernard Ahon, Ousmane Diallo & Daniel Pouakouyou May 2020 ©JNCC, Peterborough 2020 ISSN 0963-8091 For further information please contact: CITES Unit Joint Nature Conservation Committee Monkstone House City Road Peterborough PE1 1YJ United Kingdom www.jncc.gov.uk This report should be cited as: Rondeau, G., Condeé, M.M., Ahon, B., Diallo, O. & Pouakouyou, D. 2020. Survey of the occurrence and relative abundance of raptors in Guinea subject to international trade: Addendum – raptor distribution maps. JNCC Report No.412 Addendum. JNCC, Peterborough, ISSN 0963-8091. JNCC EQA Statement: This report is compliant with the JNCC Evidence Quality Assurance Policy http://jncc.Defra.gov.uk/default.aspx?page=6675. Please Note: Report 412 was originally published in 2008 and is available here. This Annex of distribution maps was produced at that time but was not published. Contents 1 Background ................................................................................................................. 1 2 References ................................................................................................................... 1 3 Raptor distribution maps ............................................................................................ 2 Pandion haliaetus - Osprey ................................................................................................... -

A Case of Cooperative Hunting by a Pair of Northern Goshawks
Acta zoologica cracoviensia, 63(2) 2020 e-ISSN 2300-0163 Kraków, 2020 https://doi.org/10.3409/azc.63.04 http://www.isez.pan.krakow.pl/en/acta-zoologica.html A case of cooperative hunting by a pair of northern goshawks Bart³omiej KUSAL and £ukasz KAJTOCH Received: 14 April 2020. Accepted: 18 September 2020. Available online: 9 October 2020. Issue online: 30 December 2020. Short communication KUSAL B., KAJTOCH £. 2020. A case of cooperative hunting by a pair of northern goshawks. Acta zool. cracov., 63(2): 21-22. Abstract. Cooperative hunting is a rare strategy in raptors, although it has been widely described in Falconidae and in some species of Accipitridae. Records about synchronous hunting in the member of the genus Accipiter are occasional. Here we describe a case of the cooperative hunting of two northern goshawks, A. gentilis, of a pigeon, Columba sp., observed in southern Poland in 2020. This exemplary behavior could be either exceptional, or cooperative hunting is an overlooked phenomenon in goshawk biology. Key words: Accipiter gentilis, Accipitridae, Columba, tandem hunting, prey. *£ukasz KAJTOCH, Bart³omiej KUSAL, Institute of Systematics and Evolution of Animals Polish Academy of Sciences, S³awkowska 17, 31-016, Kraków, Poland. E-mail: [email protected] I. INTRODUCTION and Melierax sp.). Some other birds of prey hunt in groups, but each individual preys on its own e.g. os- Cooperative hunting is a strategy that is not com- preys (Pandion haliaetus L.) and kites (Elanoides for- mon in wild animals (PACKER &RUTTAN 1988). In ficatus L., Ictinia mississippiensis (Wilson, 1811), some animal groups, cooperative hunting involves Rostrhamus sociabilis (Vieillot, 1817)). -

Disaggregation of Bird Families Listed on Cms Appendix Ii
Convention on the Conservation of Migratory Species of Wild Animals 2nd Meeting of the Sessional Committee of the CMS Scientific Council (ScC-SC2) Bonn, Germany, 10 – 14 July 2017 UNEP/CMS/ScC-SC2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II (Prepared by the Appointed Councillors for Birds) Summary: The first meeting of the Sessional Committee of the Scientific Council identified the adoption of a new standard reference for avian taxonomy as an opportunity to disaggregate the higher-level taxa listed on Appendix II and to identify those that are considered to be migratory species and that have an unfavourable conservation status. The current paper presents an initial analysis of the higher-level disaggregation using the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World Volumes 1 and 2 taxonomy, and identifies the challenges in completing the analysis to identify all of the migratory species and the corresponding Range States. The document has been prepared by the COP Appointed Scientific Councilors for Birds. This is a supplementary paper to COP document UNEP/CMS/COP12/Doc.25.3 on Taxonomy and Nomenclature UNEP/CMS/ScC-Sc2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II 1. Through Resolution 11.19, the Conference of Parties adopted as the standard reference for bird taxonomy and nomenclature for Non-Passerine species the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World, Volume 1: Non-Passerines, by Josep del Hoyo and Nigel J. Collar (2014); 2. -

Chromosome Painting in Three Species of Buteoninae: a Cytogenetic Signature Reinforces the Monophyly of South American Species
Chromosome Painting in Three Species of Buteoninae: A Cytogenetic Signature Reinforces the Monophyly of South American Species Edivaldo Herculano C. de Oliveira1,2,3*, Marcella Mergulha˜o Tagliarini4, Michelly S. dos Santos5, Patricia C. M. O’Brien3, Malcolm A. Ferguson-Smith3 1 Laborato´rio de Cultura de Tecidos e Citogene´tica, SAMAM, Instituto Evandro Chagas, Ananindeua, PA, Brazil, 2 Faculdade de Cieˆncias Exatas e Naturais, ICEN, Universidade Federal do Para´, Bele´m, PA, Brazil, 3 Cambridge Resource Centre for Comparative Genomics, Cambridge, United Kingdom, 4 Programa de Po´s Graduac¸a˜oem Neurocieˆncias e Biologia Celular, ICB, Universidade Federal do Para´, Bele´m, PA, Brazil, 5 PIBIC – Universidade Federal do Para´, Bele´m, PA, Brazil Abstract Buteoninae (Falconiformes, Accipitridae) consist of the widely distributed genus Buteo, and several closely related species in a group called ‘‘sub-buteonine hawks’’, such as Buteogallus, Parabuteo, Asturina, Leucopternis and Busarellus, with unsolved phylogenetic relationships. Diploid number ranges between 2n = 66 and 2n = 68. Only one species, L. albicollis had its karyotype analyzed by molecular cytogenetics. The aim of this study was to present chromosomal analysis of three species of Buteoninae: Rupornis magnirostris, Asturina nitida and Buteogallus meridionallis using fluorescence in situ hybridization (FISH) experiments with telomeric and rDNA probes, as well as whole chromosome probes derived from Gallus gallus and Leucopternis albicollis. The three species analyzed herein showed similar karyotypes, with 2n = 68. Telomeric probes showed some interstitial telomeric sequences, which could be resulted by fusion processes occurred in the chromosomal evolution of the group, including the one found in the tassociation GGA1p/GGA6. -

Environmental Sensitivity Index Guidelines Version 2.0
NOAA Technical Memorandum NOS ORCA 115 Environmental Sensitivity Index Guidelines Version 2.0 October 1997 Seattle, Washington noaa NATIONAL OCEANIC AND ATMOSPHERIC ADMINISTRATION National Ocean Service Office of Ocean Resources Conservation and Assessment National Ocean Service National Oceanic and Atmospheric Administration U.S. Department of Commerce The Office of Ocean Resources Conservation and Assessment (ORCA) provides decisionmakers comprehensive, scientific information on characteristics of the oceans, coastal areas, and estuaries of the United States of America. The information ranges from strategic, national assessments of coastal and estuarine environmental quality to real-time information for navigation or hazardous materials spill response. Through its National Status and Trends (NS&T) Program, ORCA uses uniform techniques to monitor toxic chemical contamination of bottom-feeding fish, mussels and oysters, and sediments at about 300 locations throughout the United States. A related NS&T Program of directed research examines the relationships between contaminant exposure and indicators of biological responses in fish and shellfish. Through the Hazardous Materials Response and Assessment Division (HAZMAT) Scientific Support Coordination program, ORCA provides critical scientific support for planning and responding to spills of oil or hazardous materials into coastal environments. Technical guidance includes spill trajectory predictions, chemical hazard analyses, and assessments of the sensitivity of marine and estuarine environments to spills. To fulfill the responsibilities of the Secretary of Commerce as a trustee for living marine resources, HAZMAT’s Coastal Resource Coordination program provides technical support to the U.S. Environmental Protection Agency during all phases of the remedial process to protect the environment and restore natural resources at hundreds of waste sites each year. -

Tinamiformes – Falconiformes
LIST OF THE 2,008 BIRD SPECIES (WITH SCIENTIFIC AND ENGLISH NAMES) KNOWN FROM THE A.O.U. CHECK-LIST AREA. Notes: "(A)" = accidental/casualin A.O.U. area; "(H)" -- recordedin A.O.U. area only from Hawaii; "(I)" = introducedinto A.O.U. area; "(N)" = has not bred in A.O.U. area but occursregularly as nonbreedingvisitor; "?" precedingname = extinct. TINAMIFORMES TINAMIDAE Tinamus major Great Tinamou. Nothocercusbonapartei Highland Tinamou. Crypturellus soui Little Tinamou. Crypturelluscinnamomeus Thicket Tinamou. Crypturellusboucardi Slaty-breastedTinamou. Crypturellus kerriae Choco Tinamou. GAVIIFORMES GAVIIDAE Gavia stellata Red-throated Loon. Gavia arctica Arctic Loon. Gavia pacifica Pacific Loon. Gavia immer Common Loon. Gavia adamsii Yellow-billed Loon. PODICIPEDIFORMES PODICIPEDIDAE Tachybaptusdominicus Least Grebe. Podilymbuspodiceps Pied-billed Grebe. ?Podilymbusgigas Atitlan Grebe. Podicepsauritus Horned Grebe. Podicepsgrisegena Red-neckedGrebe. Podicepsnigricollis Eared Grebe. Aechmophorusoccidentalis Western Grebe. Aechmophorusclarkii Clark's Grebe. PROCELLARIIFORMES DIOMEDEIDAE Thalassarchechlororhynchos Yellow-nosed Albatross. (A) Thalassarchecauta Shy Albatross.(A) Thalassarchemelanophris Black-browed Albatross. (A) Phoebetriapalpebrata Light-mantled Albatross. (A) Diomedea exulans WanderingAlbatross. (A) Phoebastriaimmutabilis Laysan Albatross. Phoebastrianigripes Black-lootedAlbatross. Phoebastriaalbatrus Short-tailedAlbatross. (N) PROCELLARIIDAE Fulmarus glacialis Northern Fulmar. Pterodroma neglecta KermadecPetrel. (A) Pterodroma -

Systematics and Conservation of the Hook-Billed Kite Including the Island Taxa from Cuba and Grenada J
Animal Conservation. Print ISSN 1367-9430 Systematics and conservation of the hook-billed kite including the island taxa from Cuba and Grenada J. A. Johnson1,2, R. Thorstrom1 & D. P. Mindell2 1 The Peregrine Fund, Boise, ID, USA 2 Department of Ecology & Evolutionary Biology, University of Michigan Museum of Zoology, Ann Arbor, MI, USA Keywords Abstract phylogenetics; coalescence; divergence; conservation; cryptic species; Chondrohierax. Taxonomic uncertainties within the genus Chondrohierax stem from the high degree of variation in bill size and plumage coloration throughout the geographic Correspondence range of the single recognized species, hook-billed kite Chondrohierax uncinatus. Jeff A. Johnson, Department of Ecology & These uncertainties impede conservation efforts as local populations have declined Evolutionary Biology, University of Michigan throughout much of its geographic range from the Neotropics in Central America Museum of Zoology, 1109 Geddes Avenue, to northern Argentina and Paraguay, including two island populations on Cuba Ann Arbor, MI 48109, USA. and Grenada, and it is not known whether barriers to dispersal exist between any Email: [email protected] of these areas. Here, we present mitochondrial DNA (mtDNA; cytochrome B and NADH dehydrogenase subunit 2) phylogenetic analyses of Chondrohierax, with Received 2 December 2006; accepted particular emphasis on the two island taxa (from Cuba, Chondrohierax uncinatus 20 April 2007 wilsonii and from Grenada, Chondrohierax uncinatus mirus). The mtDNA phylo- genetic results suggest that hook-billed kites on both islands are unique; however, doi:10.1111/j.1469-1795.2007.00118.x the Cuban kite has much greater divergence estimates (1.8–2.0% corrected sequence divergence) when compared with the mainland populations than does the Grenada hook-billed kite (0.1–0.3%). -

Spizaetus Boletin De La Red De Rapaces Neotropicales
SPIZAETUS BOLETIN DE LA RED DE RAPACES NEOTROPICALES NÚMERO 27 JUNIO 2019 BUTEO PLATYPTERUS EN COSTA RICA MILVAGO CHIMACHIMA EN COSTA RICA ASIO STYGIUS EN COLOMBIA BUTEO BRACHYURUS EN MÉXICO FALCO FEMORALIS EN MÉXICO SPIZAETUS BOLETIN DE LA RRN Número 27 © Junio 2019 Edición en Español, ISSN 2157-8966 Foto de la Portada: Juvenil de Gavilán Aludo (Buteo platypterus) en Costa Rica © Víctor Acosta-Chaves Traductores/Editores: Laura Andréa Lindenmeyer de Sousa & Marta Curti Diseño Gráfico: Marta Curti Spizaetus: Bolettin de la Red de Rapaces Neotropicales © Junio 2019 www.neotropicalraptors.org Este boletín puede ser reproducido, descargado y distribuido por fines no comerciales. Para volvera publicar cualquier artículo que figuran en este documento, por favor póngase en contacto con los autores correspondientes Contenido Singular comportamiento del caracara cabecigualdo (MILVAGO CHIMACHIMA: Falconidae) en el sur de Costa Rica José Manuel Mora & Estefanía González ............................................................................2 Nuevas localidades en Colombia del Búho Negruzco (ASIO STYGIUS) Elvis Felipe Quintero Quintero, Jeyson Sanabria-Mejía, Angélica Magaly Sogamoso Hernández & Sergio Chaparro-Herrera .................................................................................................................9 Un caso de agresión de BUTEO BRACHYURUS contra PSEUDASTUR ALBICOLLIS (Accipi- triformes) en el sur de Huimanguillo, Tabasco, México Saúl Sánchez Soto .......................................................................................................13 -

The Gambia: a Taste of Africa, November 2017
Tropical Birding - Trip Report The Gambia: A Taste of Africa, November 2017 A Tropical Birding “Chilled” SET DEPARTURE tour The Gambia A Taste of Africa Just Six Hours Away From The UK November 2017 TOUR LEADERS: Alan Davies and Iain Campbell Report by Alan Davies Photos by Iain Campbell Egyptian Plover. The main target for most people on the tour www.tropicalbirding.com +1-409-515-9110 [email protected] p.1 Tropical Birding - Trip Report The Gambia: A Taste of Africa, November 2017 Red-throated Bee-eaters We arrived in the capital of The Gambia, Banjul, early evening just as the light was fading. Our flight in from the UK was delayed so no time for any real birding on this first day of our “Chilled Birding Tour”. Our local guide Tijan and our ground crew met us at the airport. We piled into Tijan’s well used minibus as Little Swifts and Yellow-billed Kites flew above us. A short drive took us to our lovely small boutique hotel complete with pool and lovely private gardens, we were going to enjoy staying here. Having settled in we all met up for a pre-dinner drink in the warmth of an African evening. The food was delicious, and we chatted excitedly about the birds that lay ahead on this nine- day trip to The Gambia, the first time in West Africa for all our guests. At first light we were exploring the gardens of the hotel and enjoying the warmth after leaving the chilly UK behind. Both Red-eyed and Laughing Doves were easy to see and a flash of colour announced the arrival of our first Beautiful Sunbird, this tiny gem certainly lived up to its name! A bird flew in landing in a fig tree and again our jaws dropped, a Yellow-crowned Gonolek what a beauty! Shocking red below, black above with a daffodil yellow crown, we were loving Gambian birds already. -

Volume 29 Number 1 April 2011
BOOBOOK JOURNAL OF THE AUSTRALASIAN RAPTOR ASSOCIATION Volume 29 Number 1 April 2011 ARA CONTACTS President: Victor Hurley 0427 238 898 [email protected] Secretary Nick Mooney 0427 826 922 [email protected] Treasurer VACANT Webmaster VACANT Editor, Boobook Dr Stephen Debus 02 6772 1710 (ah) [email protected] Boobook production Hugo Phillipps Area Representatives: ACT Mr Jerry Olsen [email protected] NSW Dr Rod Kavanagh [email protected] NT Mr Ray Chatto [email protected] Qld Mr Stacey McLean [email protected] SA Mr Ian Falkenberg [email protected] WA Mr Jonny Schoenjahn [email protected] Tas Mr Nick Mooney [email protected] Vic Mr David Whelan [email protected] New Zealand VACANT PNG/Indonesia Dr David Bishop [email protected] Other BOPWatch liaison Victor Hurley [email protected] Editor, Circus Victor Hurley Captive raptor advisor Michelle Manhal 0418 387 424 [email protected] Education advisor Greg Czechura 07 3840 7642 (bh) [email protected] Raptor management Nick Mooney 0427 826 922 [email protected] advisor Membership enquiries Membership Officer, Birds Australia, Suite 2-05, 60 Leicester Street, Carlton, Vic. 3053 Ph. 1300 730 075, [email protected] Annual subscription $A30 single membership, $A35 family and $A45 for institutions, due on 1 January. Bankcard and MasterCard can be debited by prior arrangement. Website: www.birdsaustralia.com.au/ara The aims of the Association are the study, conservation and management of diurnal and nocturnal raptors of the Australasian Faunal Region. -

Uncorrected Proof
Policy and Practice UNCORRECTED PROOF BBarney_C011.inddarney_C011.indd 117979 99/13/2008/13/2008 44:11:24:11:24 PPMM UNCORRECTED PROOF BBarney_C011.inddarney_C011.indd 118080 99/13/2008/13/2008 44:11:24:11:24 PPMM Copy edited by Richard Beatty 11 Conservation Values from Falconry Robert E. Kenward Anatrack Ltd and IUCN-SSC European Sustainable Use Specialist Group, Wareham, UK Introduction Falconry is a type of recreational hunting. This chapter considers the conser- vation issues surrounding this practice. It provides a historical background and then discusses how falconry’s role in conservation has developed and how it could grow in the future. Falconry, as defi ned by the International Association for Falconry and Conservation of Birds of Prey (IAF), is the hunting art of taking quarry in its natural state and habitat with birds of prey. Species commonly used for hunt- ing include eagles of the genera Aquila and Hieraëtus, other ‘broad-winged’ members of the Accipitrinae including the more aggressive buzzards and their relatives, ‘short-winged’ hawks of the genus Accipiter and ‘long-winged’ falcons (genus Falco). Falconers occur in more than 60 countries worldwide, mostly in North America, the Middle East, Europe, Central Asia, Japan and southern Africa. Of these countries, 48 are members of the IAF. In the European Union falconry Recreational Hunting, Conservation and Rural Livelihoods: Science and Practice, 1st edition. Edited by B. Dickson, J. Hutton and B. Adams. © 2009 Blackwell Publishing, UNCORRECTEDISBN 978-1-4051-6785-7 (pb) and 978-1-4051-9142-5 (hb). PROOF BBarney_C011.inddarney_C011.indd 118181 99/13/2008/13/2008 44:11:24:11:24 PPMM Copy edited by Richard Beatty 182 ROBERT E. -
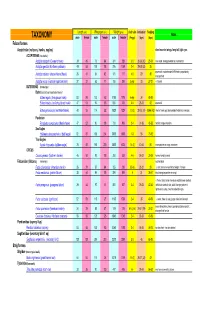
Avian Taxonomy
Length (cm) Wing span (cm) Weight (gms) cluch size incubation fledging Notes TAXONOMY male female male female male female (# eggs) (days) (days) Falconiformes Accipitridae (vultures, hawks, eagles) short rounded wings; long tail; light eyes ACCIPITRINAE (true hawks) Accipiter cooperii (Cooper's hawk) 39 45 73 84 341 528 3-5 30-36 (30) 25-34 crow sized; strongly banded tail; rounded tail Accipiter gentilis (Northern goshawk) 49 58 101 108 816 1059 2-4 28-38 (33) 35 square tail; most abundant NAM hawk; proportionaly 26 31 54 62 101 177 4-5 29 30 Accipiter striatus (sharp-shinned hawk) strongest foot Accipiter nisus (Eurasian sparrowhawk ) 37 37 62 77 150 290 5-Apr 33 27-31 Musket BUTEONINAE (broadwings) Buteo (buzzards or broad tailed hawks) Buteo regalis (ferruginous hawk) 53 59 132 143 1180 1578 6-Apr 34 45-50 Buteo lineatus (red-shouldered hawk) 47 53 96 105 550 700 3-4 28-33 42 square tail Buteo jamaicensis (red-tailed hawk) 48 55 114 122 1028 1224 1-3 (3) 28-35 (34) 42-46 (42) (Harlan' hawk spp); dark patagial featehres: immature; Parabuteo Parabuteo cuncincutus (Harris hawk) 47 52 90 108 710 890 2-4 31-36 45-50 reddish orange shoulders Sea Eagles Haliaeetus leucocephalus (bald eagle) 82 87 185 244 3000 6300 1-3 35 70-92 True Eagles Aquila chrysaetos (golden eagle) 78 82 185 220 3000 6125 1-4 (2) 40-45 50 white patches on wings: immature; CIRCUS Circus cyaneus (Northern harrier) 46 50 93 108 350 530 4-6 26-32 30-35 hovers; hunts by sound Falconidae (falcons) (longwings) notched beak Falco columbarius (American merlin) 26 29 57 64